五列木天然群体的表型变异
2013-12-27李芳东王海军廖绍波罗水兴刘东蔚
陈 勇,李芳东 ,王海军 ,孙 冰,廖绍波 ,罗水兴 ,蔡 刚 ,刘东蔚
(1.中国林业科学研究院 热带林业研究所,广东 广州 510520;2.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;3.深圳市公园管理中心,广东 深圳 518000)
五列木天然群体的表型变异
陈 勇1,李芳东2,王海军3,孙 冰1,廖绍波1,罗水兴1,蔡 刚1,刘东蔚1
(1.中国林业科学研究院 热带林业研究所,广东 广州 510520;2.中国林业科学研究院 经济林研究开发中心,河南 郑州 450003;3.深圳市公园管理中心,广东 深圳 518000)
以粤、琼两省天然分布的五列木Pentaphylax euryoides Gardn. et Champ.为研究对象,调查7个群体138个单株的7个表型性状指标,运用变异系数和单因子方差分析研究群体间和群体内的表型变异,应用相关分析研究各表型性状间的相关性以及各表型性状与地理因子间的相关性,并且运用聚类分析进行群体分类。结果表明:除群体2外,其他群体内单株间各表型性状的差异均达到显著或极显著水平。7个天然群体间叶长、基宽距、叶柄长和叶基角的差异均达到显著或极显著水平。侧脉数、叶宽、叶长宽比、基宽距、叶柄长与叶长呈显著正相关,侧脉数、叶长、基宽距、叶柄长、叶基角与叶宽呈显著正相关,叶宽和叶柄长与叶基角成显著正相关,叶长宽比与叶基角呈显著负相关。叶长、叶宽、基宽距、叶柄长以及叶基角都与经度、纬度呈极显著的正相关,叶柄长和叶基角与海拔呈极显著的正相关,基宽距与海拔呈显著相关性。通过聚类分析可将7个五列木群体分为2类:1、2、3为一类;4、5、6和7为一类。
五列木;叶片特征;地理分布;表型变异
生物的形态特征是遗传和环境综合作用的结果,其变异有其自己的遗传基础[1]。利用表型性状来研究遗传变异是简单易行且快速的方法。通过天然群体的表型变异研究,可以了解类群遗传变异的大小,同时表型变异研究是人工驯化和遗传育种研究的基础,是研究群体适应性的有效途径之一[2]。因此,目前还有许多学者应用表型性状来揭示天然群体遗传变异及其格局[3-12]。
五列木Pentaphylax euryoides Gardn. et Champ.属五列木科Pentaphylacaceae五列木属Pentaphylax,为孤属近孤种植物。天然分布于云南、贵州、广西、广东、湖南、江西、福建,木材结构很细致而均匀、纹理通直、材质稍硬、稍重,但加工容易,可供建筑、家具或农具用材。五列木树体优美、树冠浓荫苍绿,春季幼叶为嫩红色,远望去红艳夺目,具有较高的观赏价值,可作优良观赏树种。有关五列木的研究比较少,主要集中在五列木种群结构[17]和园林应用[18-19]方面,而有关五列木叶片形态特征方面尚未报道,因此笔者对分布于中国广东、海南省的7个五列木天然群体叶片形态特征进行观测,研究其叶片表型变异及其地理格局,以期为五列木的资源保护、良种选育以及城市美化提供科学依据。
1 材料与方法
1.1 数据来源
于2010年7~10月和2011年8~11月调查了五列木在海南和广东的分布范围。共收集到7个天然群体共138个单株的五列木标准枝。各调查地点五列木天然林的基本情况见表1。
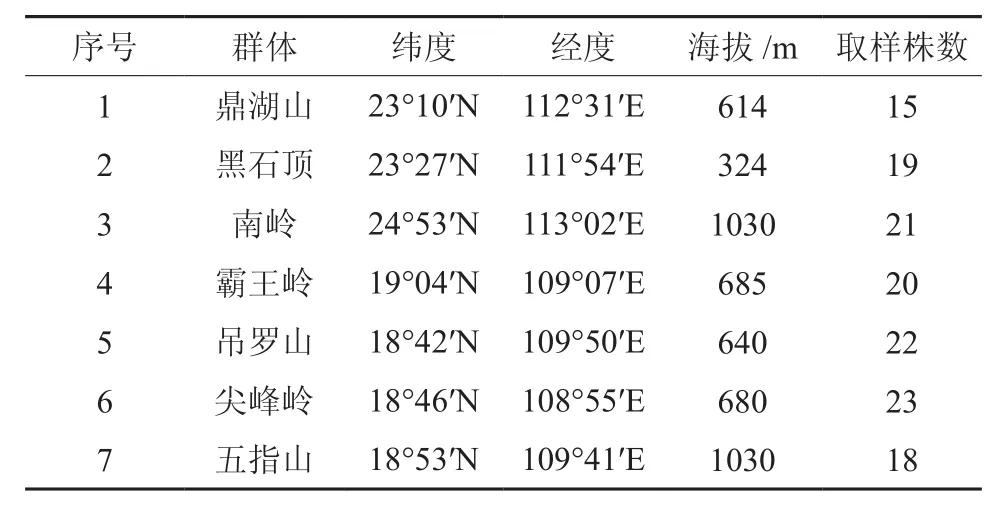
表1 广东和海南省五列木取样居群的基本概况Table 1 Basic situation of sampling P. euryoides populations in Guangdong and Hainan
对每个调查林分,原则上选取不少于20株母树,母株之间距离保持在50 m以上。要求母树生长正常,没有病虫害。取样部位是植株中部朝南方向,采集当年生顶端枝叶。
1.2 表型测定
在每株采种母株随机选取6~10片发育完满的完整叶片,测量其侧脉数、叶长、叶宽、叶基宽距(叶基至叶片最宽处距离)、叶柄长、叶基角。应用量角器、直尺等常规测量工具测量(读取2位数)。
1.3 数据分析
用叶片的长宽比,作为叶片的形状指数。采用SPSS 11.5软件对数据进行分析,包括对7个群体的7个叶片表型性状进行差分分析和多重比较,分析五列木群体的叶片表型变异特征;利用相关分析法计算叶片表型性状间的相关性及其叶片表型性状与地理间的相关性,探讨五列木表型变异的地理格局;根据群体表型性状,应用SPSS 11.5软件构建群体聚类图[4]。
2 结果与分析
2.1 五列木表型性状的变异特征
2.1.1 群体内的变异
变异系数表示性状的离散程度,反映形态多样性的丰富程度[5],因此以变异系数反映五列木叶片表型的变异程度。7个五列木群体除了群体2外,其他群体内单株间叶片表型性状的差异均达到显著或极显著水平。从表2可以看出,群体内各性状的变异大小因性状而异。其中基宽距的变异最小,平均变异系数为11.41%;叶柄长的变异最大,平均变异系数为17.29%。综合分析各性状可以看出,群体1的变异最丰富,其次是群体3,群体2的变异最小。
2.1.2 群体间的变异
从单因素方差分析结果(见表2)可以看出,7个五列木天然群体间各单株间叶长、基宽距、叶柄长和叶基角的差异均达到显著或极显著水平。在群体平均值的基础上求得群体间各表型性状的变异系数。比较群体间各表型的变异系数,基宽距的变异最小,变异系数为13.82%;叶柄长的变异最大,变异系数为40.36%。
2.2 叶片表型性状间的相关性
叶片表型性状间的相关性分析(表3)表明,侧脉数、叶宽、叶长宽比、基宽距、叶柄长与叶长呈显著正相关,侧脉数、叶长、基宽距、叶柄长、叶基角与叶宽呈显著正相关,叶宽和叶柄长与叶基角成显著正相关,叶长宽比与叶基角呈显著负相关。

表2 7个五列木天然群体内和群体间的叶片表型变异Table 2 Leaves phenotypic variations of among and within 7 P. euryoides natural populations

表3 五列木7个叶片性状间的相关分析†Table 3 Correlation analysis of 7 P. euryoides among 7 leaf characters
2.3 叶片表型性状的地理变异
树木叶片表型的地理变异比较复杂,不同物种其变异不同。由表4可知,叶长、叶宽、基宽距、叶柄长以及叶基角都与经度、纬度呈极显著的正相关,即越往东北,叶长、叶宽、基宽距、叶柄长以及叶基角越大;叶柄长和叶基角与海拔呈极显著的正相关,基宽距与海拔呈显著正相关性,即海拔越高,叶柄长、叶基角以及基宽距越大。以上不同树种叶片表型性状的地理变异,可能与其分布范围、生长环境以及干扰程度等因素有关。
2.4 群体分类
依据叶片性状对7个五列木天然群体进行聚类分析(图1),可将其分为2类:1、2、3为一类,处于广东省;4、5、6和7为一类,处于海南省。地理上相连的群体聚在一起。这说明利用五列木群体表型性状进行群体的初步区划是可行的[13-15]。
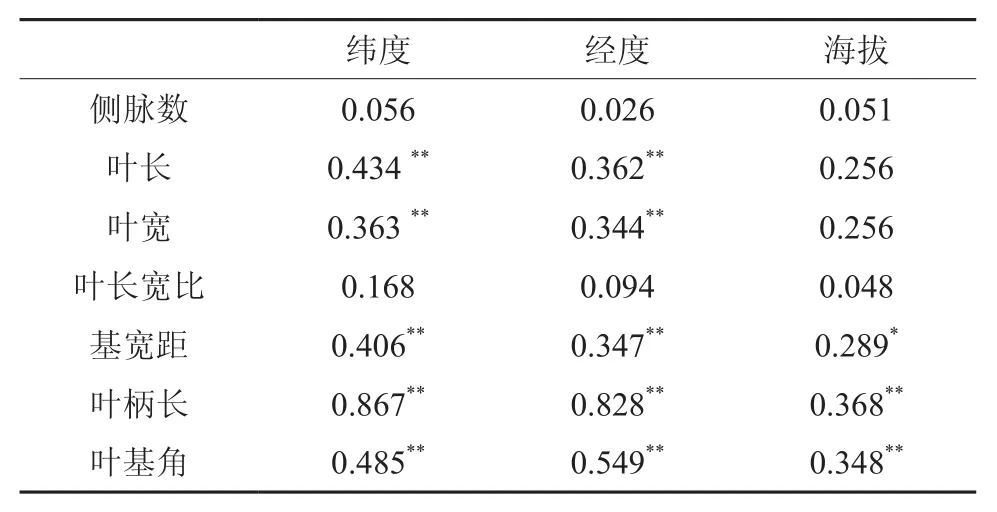
表4 五列木叶片表型性状与地理因子的相关性分析Table 4 Correlation analysis of P. euryoides between 7 leaf characters and geographical factors

图1 7个五列木天然群体聚类分析Fig.1 Cluster analysis of 7 natural populations of P. euryoides
3 结论与讨论
对五列木7个天然群体表型性状的分析表明,除群体2外,其他群体内单株间各表型性状的差异均达到显著或极显著水平。7个天然群体间叶长、基宽距、叶柄长和叶基角的差异均达到显著或极显著水平。这些变异是自身遗传因素和环境因素共同作用的结果。这种多层次的变异为种质资源和生物多样性保护提供了物质基础,同时也表明了生物多样性保护任务的艰巨性。表型性状变异一般具有适应上的意义,如叶柄长度影响树汁传导的同时影响叶片的空间分布,从而对光的截留具有重要作用[16]。
树木叶片表型的地理变异比较复杂,不同物种其变异不同。本研究中,五列木叶长、叶宽、基宽距、叶柄长以及叶基角都与经度、纬度呈极显著的正相关,即越往东北,叶长、叶宽、基宽距、叶柄长以及叶基角越大;叶柄长和叶基角与海拔呈极显著的正相关,基宽距与海拔呈显著正相关性,即海拔越高,叶柄长、叶基角以及基宽距越大。而罗建勋[6]研究表明,云杉针叶宽度与纬度和经度呈负相关,与海拔呈正相关,即群体针叶宽度随纬度和经度变异,从南向北,从西向东针叶宽度逐渐变小。江洪[7]认为云杉针叶宽度随海拔增高而增大,云杉针叶性状的变异与适应性有关。西南桦叶片侧脉数、基宽距都与经度、纬度呈现显著的负相关,也就是说越往东北方向,侧脉数和基宽距越小,而叶片长度与经度亦呈现显著的负相关[8]。
本研究通过聚类分析将7个五列木群体分为2类:1、2、3为一类,处于广东省;4、5、6和7为一类,处于海南省,地理上相连的群体聚在一起。这说明利用五列木群体表型性状进行群体的初步区划是可行的。而Barzdajn[15]认为表型是基因型和环境共同作用的结果,可采用表型性状进行欧洲云杉群体的鉴别,并根据挪威云杉群体球果和种鳞等表型性状将欧洲云杉20个种源进行区划。
五列木表型变异研究对开展种质资源保存、良种选育以及天然林保护和经营具有十分重要的意义。由于五列木资源收集和良种选育工作刚刚起步,各地生产部门可以就近进行单株选优,采集优树上的种子发展五列木人工林。在种质资源迁地或就地保存时,可以选择变异较为丰富的群体,如1、3两个群体。由于五列木天然林资源破坏较严重,需尽可能多地保护和经营现存的资源,对以上7个群体必需加以重点保护。
[1] C Brochmann, Pamela S Soltis, Douglas E Soltis. Recurrent formation and polyphyly of Nordic polyploids in Draba(Brassicaceae) [M]. Botanical Society of Americal, 1992, 79(6):673-688.
[2] 葛 颂, 洪德元. 遗传多样性及其检测方法[A].见:钱迎倩,马克平主编.生物多样性研究的原理与方法[C]. 北京: 中国科学技术出版社, 1994:123-140.
[3] Olfeh J J. Furnier G R, Luby J J. What data determine whether a plant texon is distinct enough to merit legal protection? A case study of Sedum integrifolium (Crassulaceae)[J]. Amer. J. Bot.,2001, 88(3): 401-410.
[4] 赵志刚, 郭俊杰, 沙 二, 等. 我国格木的地理分布与种实表型变异[J]. 植物学报, 2009, 44(3): 338-344.
[5] 李 斌, 顾万春, 卢宝明. 白皮松天然群体种实性状表型多样性研究[J]. 生物多样性, 2002,10:181-188.
[6] 罗建勋, 李晓清, 孙 鹏, 等. 云杉天然群体的表型变异[J].东北林业大学学报, 2003, 31(1): 9-11.
[7] 江 洪. 云杉种群生态学[M]. 北京:中国林业出版社,1992.
[8] 曾 杰, 郑海水, 甘四明, 等. 广西西南桦天然居群的表型变异[J]. 林业科学, 2005,41(2): 59-65.
[9] 陈 璋. 福建山樱花天然居群表型变异研究[J]. 热带亚热带植物学报, 2008, 16(1): 61-68.
[10] 王雷宏, 汤庚国. 山荆子腊叶标本表型性状变异分析[J]. 西北植物学报, 2007, 27(8): 1690-1694.
[11] 曾 杰. 植物天然居群表型变异研究的几个热点问题[J]. 广西林业科学, 2007, 36(2): 65-70.
[12] 李 梅, 韩海荣, 康峰峰, 等. 山西灵空山辽东栎种群叶性状表型变异研究[J]. 北京林业大学学报, 2005, 27(5): 10-16.
[13] Atasay H. Studies of the genetic diversity within and between the population of oriental spruce (Picea orientlis) from the characteristics of seeds and seedlings[J] . Teknik Bulten Serisi Ormancilik Arastirma Emstitusu Yayinlari, 1996, 261:86.
[14] Barzdajn W. Intra population diversity in spruce (Picea abies(L.)Karst.) in the mountains[J]. Sylwan, 1996, 140:3, 39-46.
[15] Barzdajn W. An assessment of diagnostic value of morphological traits of Norway spruce (Picea abies (L.) Karst.) cones for discrimination of spruce provenances[J]. Sylwan, 1996,140:9,61-75.
[16] Bacilieri R., Ducousso A.& Kremer A. Genetic, morphological,ecological and phonological differentiation between Quercus petraea (Matt.) Liebl. and Quercus robur L. in a mixed stand of northwest of France[J]. Silvae Genetica,1995,44(1):1-10.
[17] 张桂清, 谢 强,李 锋, 等. 桂东北山地五列木种群结构与动态的初步研究[J]. 广西植物2008, 28(1): 62-66.
[18] 黄文韬,廖菊阳. 五列木的园林应用初探[J]. 湖南林业科技,2009, 36(5): 23-25.
[19] 张强英, 张启翔, 程堂仁. 榆叶梅野生群体表型多样性的研究[J]. 中南林业科技大学学报, 2012, 32(5): 155-160.
Phenotypic variation of leaves of Pentaphylax euryoides Gardn. et Champ.natural populations
CHEN Yong1, LI Fang-dong2, WANG Hai-jun3, SUN Bing1, LIAO Shao-bo1, LUO Shui-xing1, CAI Gang1, LIUDong-wei1
(1. Research Institute of Tropical Forestry, Chinese Academy of Forestry, Guangzhou 510520, Guangdong, China; 2.Non-timber Forestry Research and Development Center, Chinese Academy of Forestry, Zhengzhou 450003, Henan, China; 3. Shenzhen Park Service,Shenzhen 518000, Guangdong, China)
Seven phenotypic traits were investigated on 7 natural populations of Pentaphylax euryoides Gardn. et Champ. from Guangdong and Hainan of China, including 138 individuals. The phenotypic variations among the populations and within the populations were studied by using variation coefficients method and F-test of one-way variance analysis. The correlation among the phenotypic characters and the correlation between the phenotypic characters and geographical factors were studied. The natural population classif i cation was conducted by using clustering analysis method. The results show that within 6 populations, the differences of phenotypic characters among single plant all reached significant level or very significant level except that of population No.2.Among 7 populations, length of leaves, height of max width, length of petiole and angle of leaf base were signif i cant or quite signif i cant differences; number of lateral vein, width of leaves, leaf length/width, height of max width, length of petiole was remarkably positively correlated to length of leaves. Number of lateral vein, length of leaves, height of max width, the length of petiole was remarkably positively correlated to width of leaves. Width of leaves, length of petiole was remarkably positively correlated to angle of leaf base.Leaf length/width was remarkably negatively correlated to angle of leaf base. Among 7 phenotypic traits, length of leaves, width of leaves, height of maximum width, length of leaves to longitude, length of petiole and angle of leaf base were remarkably correlated to longitude and latitude, length of petiole and angle of leaf base to altitude, height of maximum width to altitude. Hierarchical cluster analysis revealed that these 7 populations can be divided into two groups: (1)No.1, No.2 and No.3 belong to fi rst group ; (2) No.4, No.5,No. 6 and No.7 second group.
Pentaphylax euryoides Gardn. et Champ.; leaf traits; geographic distribution; phenotypic variation
S792.99 ;S718.46
A
1673-923X(2013)02-0020-04
2012-10-16
中央级公益性科研院所基本科研业务费专项资金项目(RITFKYYW2010-12);深圳市科技计划项目“深圳市优良乡土树种选育及应用”(JSA200904031813A)
陈 勇(1978-),男,湖南人,助理研究员,博士研究生,从事城市林业方面的研究
廖绍波(1957-),男,高级工程师,主要从事城市林业方面的研究;E-mail:lshaobo@126.com
[本文编校:吴 彬]
