人工高效经营雷竹林CO2通量估算及季节变化特征
2013-12-25陈云飞周国模
陈云飞,江 洪,2,*,周国模,杨 爽,陈 健
(1.浙江农林大学浙江省森林生态系统碳循环与固碳减排重点实验室,浙江 311300;2.南京大学国际地球系统科学研究所,南京 210093)
温室气体排放使得地球生物化学循环碳收支失衡,森林生态系统在吸收CO2维持生物圈碳氧平衡过程中作用巨大,可计量的森林碳汇日益成为国际研究的热点。研究生物圈的碳循环机理和全球陆地生态系统碳收支及其对环境变化的响应已成为当今一系列大型国际计划共同关注的核心问题,科学界急切的要揭示“碳失汇”问题[1],这部分未知的碳汇有待研究。CO2和H2O通过植被的光合作用和呼吸作用在土壤-植被-大气圈空间层次上时刻进行着交换,以涡度相关技术为主体对土壤-植被-大气间的CO2/H2O和能量通量以及生态系统碳水循环的关键过程进行长期连续的观测,所获取的观测数据将被用来量化和对比分析研究区域内的生态系统碳收支与平衡特征及其对环境变化的响应[2]。目前涡度相关技术已在全球范围内广泛的应用于陆地生态系统的碳通量吸收与排放测定中,并取得了很好的成效,该方法已成为国际通量观测网(FLUXNET)的标准方法。中国陆地生态通量观测网络(ChinaFLUX)也已经利用该技术开展了广泛的观测[3-4]。
亚热带季风气候分布在北纬25°—35°亚热带大陆东岸地区,分布地区包括日本、韩国,美国、巴西的部分地区,中国的秦岭-淮河以南的区域,并以中国面积最大。在亚热带通量观测的系统还很有限,只有江西千烟州人工针叶林观测站站以及哀牢山地常绿阔叶林观测系统,两站点南北跨度大仅对两大植被类型进行了观测,而亚热带植被类型以常绿阔叶林为主,混杂有落叶林、针叶林、混交林和竹类,亚热带季风区域植被类型丰富,同时多丘陵,森林覆盖率高,有巨大的碳汇潜力和重要的碳源汇角色。竹林的通量观测正是对亚热带通量观测的补充和深入研究。
雷竹(Phyllostachysviolascens)作为经济竹类在长江以南均有种植,且种植面积日益扩大。竹林采伐期短,更新代谢快,高效经营,收获竹笋经济效益显著,同时应对气候变化,增加森林碳汇有着积极的意义。独特的管理方式和生理生态过程使其兼具森林、农田两大系统的特点,雷竹林生态系统较一般森林,作为常绿树种全年光合能力强,采伐期短更新代谢快,经济林型种植面积广大。很多研究表明,青幼年森林的碳吸收能力要强于衰老森林。北美黑云杉的采伐对碳源汇的影响[5],EOBS站点为采伐前林分年龄105a微弱碳汇(6±4)Cg·m-2·a-1, HBS75站点青幼年林型林分年龄33a为强的碳汇(143±35)Cg·m-2·a-1。因此,从生物学角度雷竹始终为青幼年的林龄,应具有较强碳汇潜力,但竹林的通量过程与固碳能力国内外文献报道很少,有待于深入研究。在亚热带地区建立长期连续观测的通量塔,对揭示竹林碳源汇过程以及影响碳平衡机制有深刻意义,对区域尺度下特色生态系统的碳平衡研究有启发指导作用。
本文以浙江省临安太湖源雷竹林生态系统观测塔的一年通量数据为依据,结合雷竹高效经营覆盖耕作科学的管理措施[6]。分析雷竹林系统CO2通量变化过程和碳收支量,及其人为干扰和环境因子对CO2通量变化的影响。
1 材料与方法
1.1 观测站点
试验区位于浙江省临安市太湖源镇,地理位置N 30°18′169″ E119°34′104″属亚热带季风气候,温暖湿润,雨热同期,气候特征春季多雨,夏湿热有梅雨期,秋气爽,冬干冷。全年降水量1600mm,年平均气温16℃,全年日照时数1900h。试验地土壤以红壤为主,海拔185m,坡向北偏东35°,坡度2—3°,雷竹林建园历史10 a。观测塔下雷竹林群落平均高度4.5m,平均胸径4cm,以2、3年生竹为主,总盖度80%,立竹密度每亩1500株,林下灌木草本很少,有竹叶糠皮覆盖,为人工经营的笋用经济林。雷竹是禾本科竹亚科刚竹属竹种,由于早春打雷即出笋而得名,实为早竹之变种。自然状态下雷竹,秆高7—11m,径达4—6cm。节间较短而均匀,幼竿密被白粉生长季以1a为1个周期,连年出笋能力强,在出笋期与笋芽分化期要求有充足降水[7]。
人工高效经营的雷竹主要措施是充足肥料供给、充足水分灌溉、钩梢剪枝、间伐老竹、覆盖增温等。其中冬季采用双层法覆盖增温是关键技术,用砻糠、稻草双层覆盖,其厚度30cm(下面铺稻草10—15cm,上面覆盖砻糠15—20cm)。自然状态下,2月下旬到3月上旬土壤温度10℃以上,竹出笋最低温度需9—10℃,最适温度20—24℃[8],有效积温200℃。4月份覆盖的稻草腐烂入土,砻糠及时移除。覆盖增温后,11月底竹林覆盖稻草砻糠保持地温10℃以上,使雷竹提前发笋12月底开始萌动。1月中下旬大量出土,恰赶春节前后上市,价格较高。雷竹林每年都要收获竹笋留养新竹,伐走等量的4a老竹,相当于间伐的管理措施,间伐量约占竹林的20%—30%,始终保持一定的立竹密度和1—3a的竹龄,这样竹林生态系统处于发育的青幼时期,其碳汇能力较森林和农田系统有明显优势。
1.2 观测仪器
观测竹林建有高20m的微气象观测塔,开路涡度相关系统的探头安装在17 m高度上,由三维超声风温仪(CAST3, Campbell Inc., USA) 和开路CO2PH2O 分析仪(Li-7500, LiCor Inc., USA) 组成,原始采样频率为10Hz,数据传输给数据采集器(CR1000, Campbell Inc., USA) 进行存储,同时根据涡度相关原理在线计算并存储30min 的CO2通量(FC)、潜热通量(LE)和显热通量(HS)等统计量。常规气象观测系统,包括3层风速(010C, met one, USA) 、3层大气温度和湿度(HMP45C,Vaisala, Helsinki, Finland),安装高度分别为1m,5m, 17m,2个SI-111红外温度分别置于1.5m和5m,用于采集地表和冠层温度,净辐射仪(CNR4,Kipp&Zonen)传感器安装高度17m,用于采集上行/下行的长波/短波辐射、净辐射的数据,此外还有土壤热通量(HFP01,Hukseflux)观测深度3cm,5cm;土壤含水量(CS616,Campbell,USA)观测深度5,50,100cm;土壤温度(109,Campbell,USA)观测深度5,50,100cm。常规气象观测系统数据采样频率为0.5 Hz,通过数据采集器(CR1000,Campbell Inc,USA) 每30 min自动记录平均风速、温度、气压、净辐射等常规气象信息。常规气象数据用于插补通量观测数据的缺失。7层CO2/H2O廓线观测系统(LI-840,Li CorInc,USA)观测CO2/H2O从土壤到植被再到大气的扩散过程,为闭路系统,采样频率为0.2Hz,同时输出30min平均值。全部观测数据保存到数据采集器的TF卡上,同时配置无线传输模块利用远程计算机加载虚拟串口通过手机信号连接数据采集器,可以实时监测传感器状态,30min数据可以实时传输。
1.3 研究方法
开路涡度系统数据采集器可在线计算虚温订正和空气密度变化订正。但是未考虑地形和仪器倾斜影响,当下垫面有倾斜度时,由于地球引力作用,顺着山坡走向大气会发生汇流、漏流现象,此时平均垂直风速并不为零。很多研究都是通过旋转风向坐标轴来计算通量[9-11]。在中尺度大气环流时,可以旋转坐标轴迫使平均垂直风速为零[12]。根据风向、仪器底座、主风向地形坡度等建立一新的坐标轴参考系统,经过坐标旋转后,净生态系统交换量即为涡度相关通量、储存项、水平和垂直方向的平流3项的和。常见的有二次坐标旋转、三次坐标旋转和平面拟合,本文采用二次坐标旋转(DR),使坐标系x轴与平均水平风方向平行,从而使平均侧向风速度和平均垂直风速度为零。
本文的观测方法采用的是涡度相关的微气象观测方法,通过测定大气中湍流运动产生的风速脉动和气体浓度脉动,计算二者协方差求解通量值[13]。
净生态系统碳交换量(NEE)主要是指生态系统中植物光合作用、植被冠层空气的碳储存和生态系统呼吸消耗的碳排放引起的生态系统碳储量变化[14]
微气象学研究以大气为对象,当CO2从大气进入到生态系统时,定义NEE符号为负;当CO2从生态系统排放到大气中时,定义NEE符号为正。生态学家研究的重点在生态系统的变化,对于通量的符号规定为:气体由大气圈进入生态系统的通量值符号为负,反之为正[15]。
净生态系统生产力(NEP),定义的符号刚好和NEE符号相反,生态系统总交换量(GEE)与生态系统总初级生产力(GPP)符号也是相反的,这样陆地和大气之间的气体交换过程中的关系,可用下列方程描述:
GEE=NEE-RE
(2)
NEE=Fc+Fs
(3)
RE=REnight+REday
(4)
式中,Fc为大气和生态系统冠层的碳通量,即涡度探头观测值,Fs为冠层内的碳储存通量(雷竹林高度不足8m年际变化可忽略);式中RE生态系统呼吸包括植物自养呼吸以及土壤微生物分解土壤有机质和凋落物呼吸通量,分为白天与夜晚计算。夜间生态系统完全为CO2排放状态,NEE数值上就等于生态系统呼吸值RE。白天的生态系统呼吸是通过拟合半小时时长的5cm土壤温度数据与夜间NEE数据,建立温度与生态系统呼吸的关系式,推算REday。根据公式(2),(4)。分解NEE可以分解成GEE和RE,进而计算出GEE。
1.4 数据校正与插补
基于涡度原理开路系统采集的10Hz原始数据,因为用红外气体仪观测的CO2气体浓度是相当于干空气的质量混合比,大气的温度、压力、湿度发生变化会引起CO2质量浓度的变化,需要根据理想气体状态方程校正气体密度为摩尔质量比。需要进行水汽校正,即WPL校正。根据垂直平均风速为零假设,做坐标轴旋转校正。以上校正应用EdiResoftwar设定参数,加载模块,计算成为30min步长数据。EdiRe software是由爱丁堡大学编写,专门处理涡度观测数据,并做相关校正。
在实际观测中由于受到降水、凝水、昆虫以及随机电信号异常等的影响,需要对通量数据进行质量控制,结合涡度相关法通量观测原理和ChinaFLUX推荐筛选标准,目前通用的涡度数据处理,有平均日变化法(MDV),查表法(LookUp-table),非线性回归法(NLR)。本文中应用非线性的经验方程,光合有效辐射数据偏差较大,插补数据的结果不理想相关性很小,因此采用平均日变化法。平均日变化法是对缺失数据用相邻几天同时刻数据的平均值进行查补。该方法首先确定平均时段的长度,研究发现白天取14d、夜间取7d的平均时间长度时偏差最小[16]。
涡度相关开路系统数据易受降水、电信号、仪器故障影响造成数据缺失或者不合理,一般来说文献报道的国际通量网络各个站点数据缺失和不合理比例在17%—50%[17]。美洲通量网(AmeriFlux)的站点白天缺失和不合理数据在20%—35%[18]。本站点白天数据有效率在80%左右;夜间湍流不充分CO2过多的沉积在林冠下部不能被探头检测到,筛选风摩擦系数较大的数据,有效数据在48%左右。表1说明除8月份数据有效率偏低外,其余各月数据有效率要高于通量观测网络系统的平均水平。
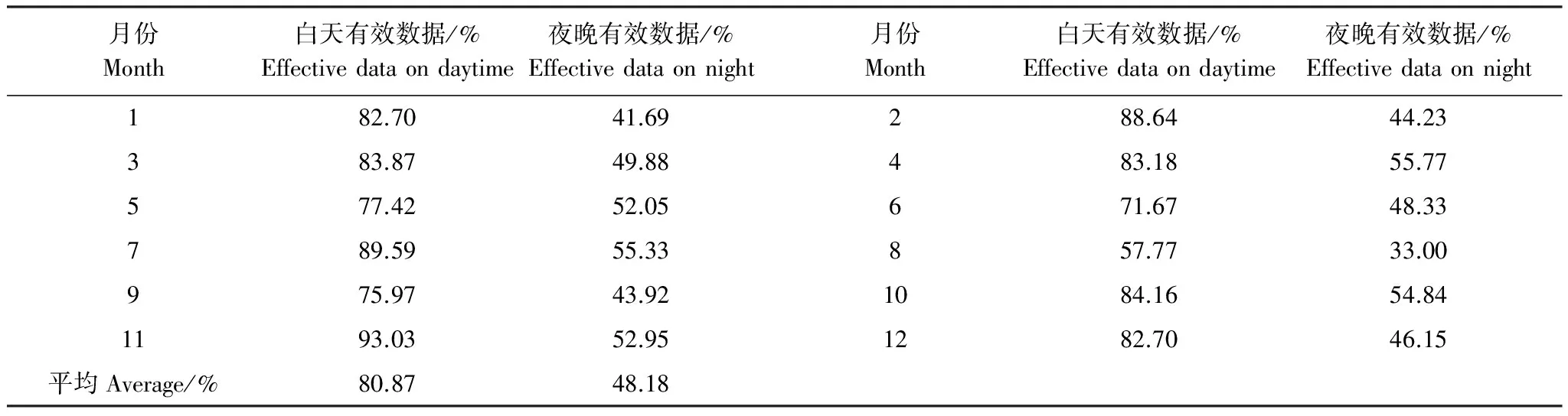
表1 各月有效数据统计
2 结果与分析
2.1 常规气象因子的季节变化
观测数据从2010年10月到2011年9月,构成全年数据。日平均最高温2011年7月8日30.21℃,最低温2011年1月17日-5.50℃。月平均气象因子见表2。全年最高气温7月,最低气温1月;光合有效辐射变化与气温相同;降雨量最大6月,最小12月,降雨与该地区往期持平,降雨量变化较气温变化年际差异大。
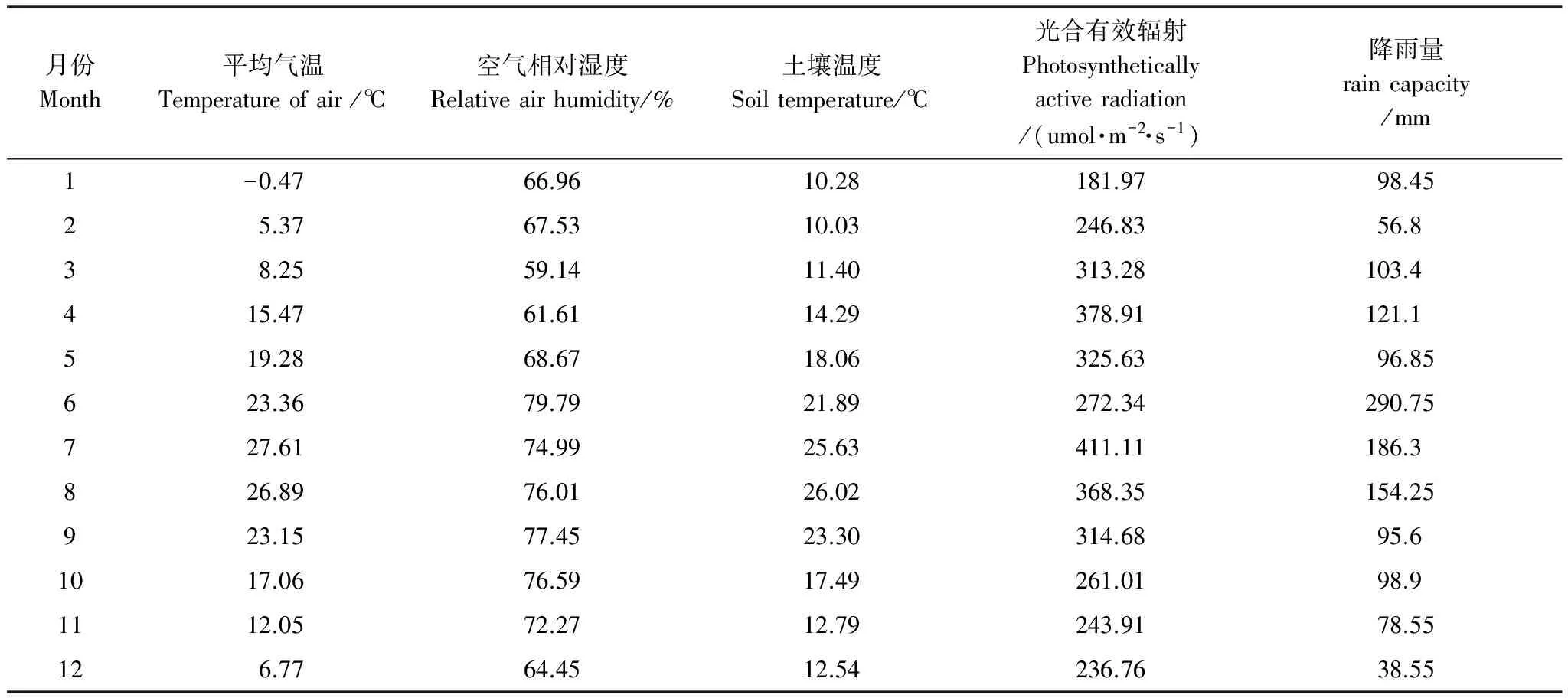
表2 各月气象因子变化
2.2 净生态系统碳交换量
从图1月每时刻平均的日进程可以看出,NEE呈“U”形变化,表现为在白天通量负值,夜间通量正值,雷竹林生态系统月尺度上的NEE符号变化时刻和最大的碳排放、碳吸收时刻都有明显的差异。各月通量符号变化,由正值转为负值在日出1—1.5h,春冬季与夏秋季相比延迟1h;由负值转为正值时刻较集中,发生在17:00—18:00。9—11月NEE由正转负发生在6:30—7:00,NEE最大负值出现在11:00前后。在12月—翌年2月NEE由正转负发生在7:30—8:00,NEE最大负值出现在12:00前后。3—8月的通量符号变化时刻和最大负值时刻介于秋季与冬季之间,NEE由正转负发生在7:00—7:30,NEE最大负值出现在11:00—11:30。
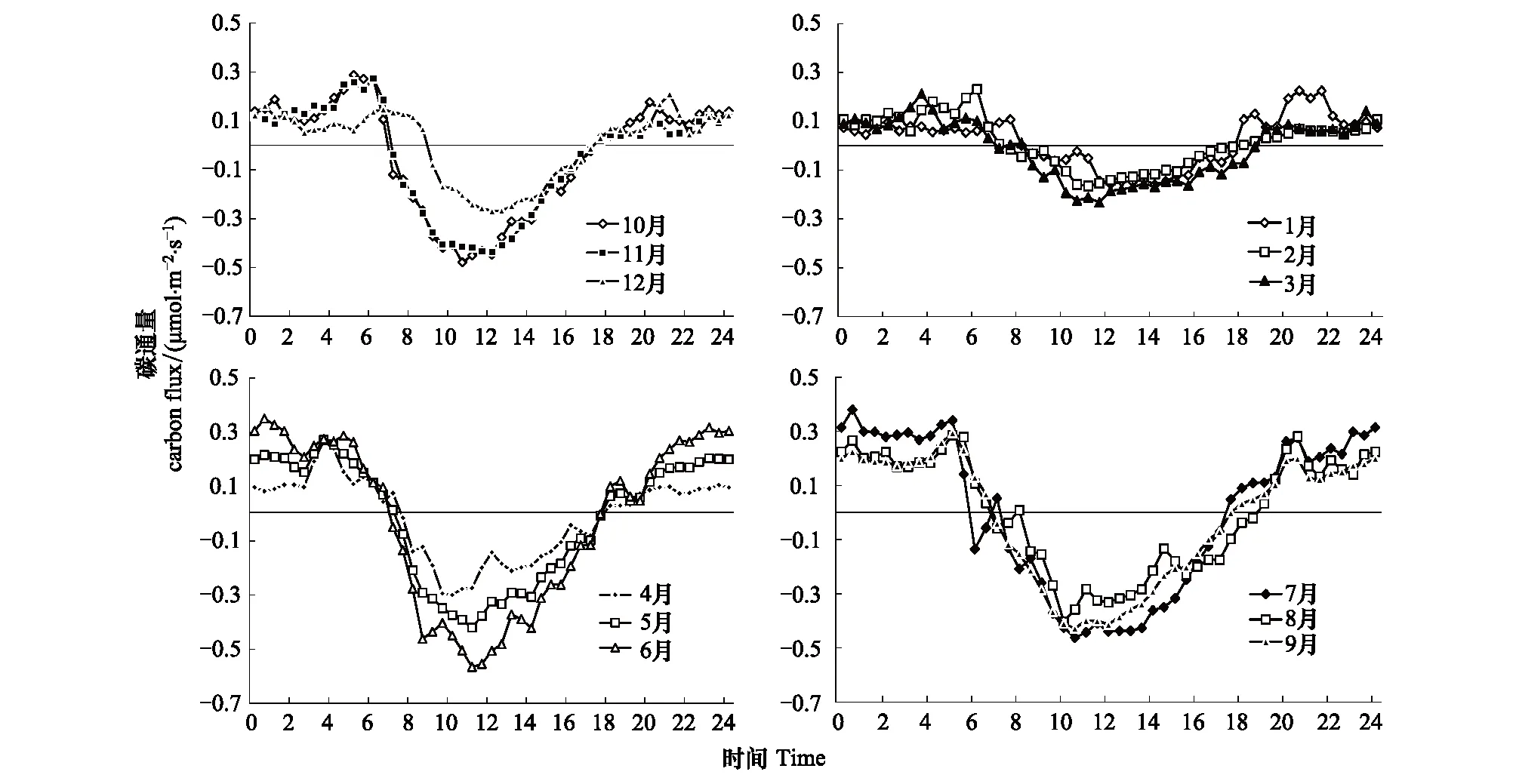
图1 CO2通量各月平均日变化
从全年NEE 变化看,年碳吸收总量126.303gC·m-2·a-1,总体是碳汇。这要小于周国模应用生物量调查法对毛竹固碳能力509gC·m-2·a-1,杉木为348gC·m-2·a-1的估算[19]。李静洁等研究江西红壤丘陵地区双季稻碳收支[20],在不同施肥措施下估算早稻田两季稻的碳汇能力为311—629gC·m-2·a-1,雷竹年固碳能力要小于水稻,同时也小于北方农田系统198—318gC·m-2·a-1 [21]年吸收能力。具体季节分析12—2月即冬季覆盖月份NEE为正,雷竹林表现为碳源,春季、夏季、秋季均为碳汇。全年碳通量,吸收量表现为双峰变化曲线,第1峰出现在6月(-21.39gCm-2·月-1);第二峰出现在11月(-35.89gC·m-2·月-1)。最高排放量1月(16.061gC·m-2·月-1)。
6月全月净积累总量要小于11月,这可能是由于亚热带6月份开始进入梅雨期,影响雷竹的光合生产能力。同纬度气候下千烟洲站点6月出现的净积累量也小于11月的净积累量[22]。7—8月受高温影响有明显的“午休”现象,CO2通量一般在10:0—11:00达到峰值,之后降低,09—11月适宜的水热条件使碳吸收持续增加可为冬季竹笋萌发积蓄有机物。7—8月份虽然有较强的初级生产力,但由于呼吸过程排放的碳也比较大,碳净吸收低于5—6月和9—11月。这在很多类型的生态系统普遍存在,在竹类系统也得到了验证。12月进入冬季开始覆盖增温,人为经营措施干扰NEE开始减小。
从图2可以看出RE呈单峰变化,在夏季较高冬季较低。图5中显示夜间NEE与土壤5cm温度的散点图,拟合曲线显示夜间NEE与土壤5cm温度成指数关系变化,相关系数0.3。7月土壤温度最高月份,RE达到峰值151.05 gC·m-2·月-1,在气温较低的春冬季呼吸较低。从季节上分析,分为冬季(12—2月),春季(3—5月),夏季(6—8月),秋季(9—11月)。各季节RE占全年总量的比例分别为16.7%,21.2%,37.0%,25.1%;各季节NEE占全年比例分别为-21.3%,26.6%,25.8%,68.9%;各季节GEE占全年比例为12.8%,21.8%,35.8%,29.6%。全年RE贡献最大为夏季,夏季生态系统呼吸是冬季的2.3倍;全年NEE贡献最大为秋季超过全年其余月份之和,而冬季为负贡献降低了全年碳汇总量,春夏季NEE贡献为年平均水平;全年GEE最大贡献为秋季,最小为夏季,变化与RE一致。
全年GEE季节变化特征明显,峰值出现在6—7月,可能与温度、水分影响有关;之后开始下降,但秋季生长明显大于春季,冬季GPP最小。根据雷竹的生长特性,竹笋出土吐叶到成竹经30—40 d,5月大量新竹开始光合生产,6月下旬到7月梅雨期,为雷竹生长提供充足的水分供给。冬季的防雪钩梢和温度影响,可能是生产力开始下降的主要影响因素。覆盖月份为12—3月,出笋是1—3月,竹笋在适宜的水热条件下爆发性生长,竹笋出土呼吸加大,同时覆盖后土壤和覆盖物分解呼吸大大超出覆盖前的土壤呼吸,有研究发现覆盖后是覆盖前的5—10倍[23],造成冬季CO2排放不降。土壤呼吸、覆盖物呼吸以及植物呼吸都土壤温度密切相关,1月温度降低,生态系统呼吸随之降低。6月GEE与RE同时上升出现峰值,新竹不断增强的光合作用吸收了一定CO2,同时生态系统呼吸达到了较大值,造成NEE不为最高值。9月GEE与RE同时开始下降时,NEE开始增加,并在11月达到全年的最高峰值。在春季NEE随GEE与RE的升高而增加,在秋季NEE随GEE与RE的下降而增加,出现两个峰值。由此可见人工高效经营下,温度是主要的影响因子,光照次之。雷竹喜湿怕涝,在受水分胁迫时人工会及时补水排涝,水分影响这里先不考虑。
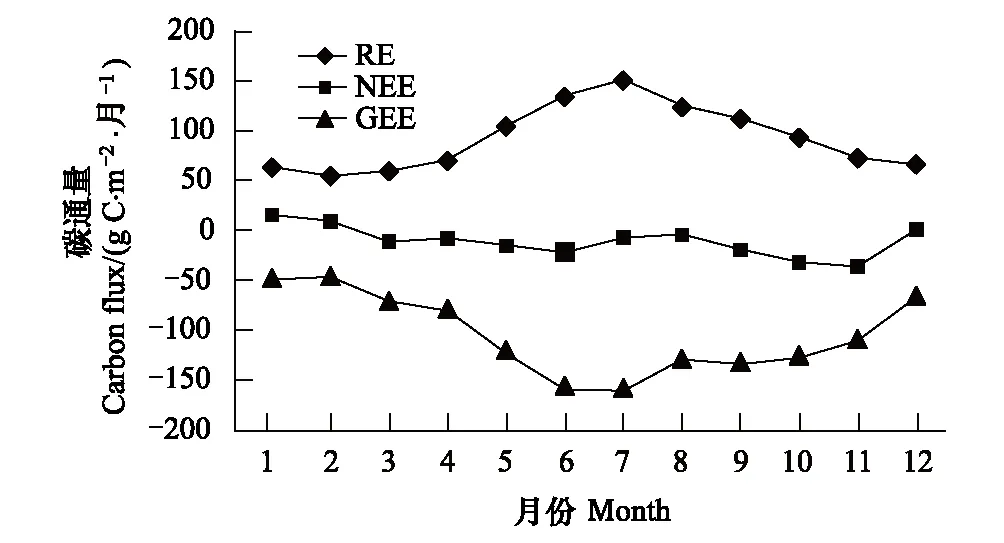
图2 全年各月累积净生态系统CO2交换量(NEE)、生态系统呼吸(RE)和总生态系统CO2交换量(GEE)变化过程
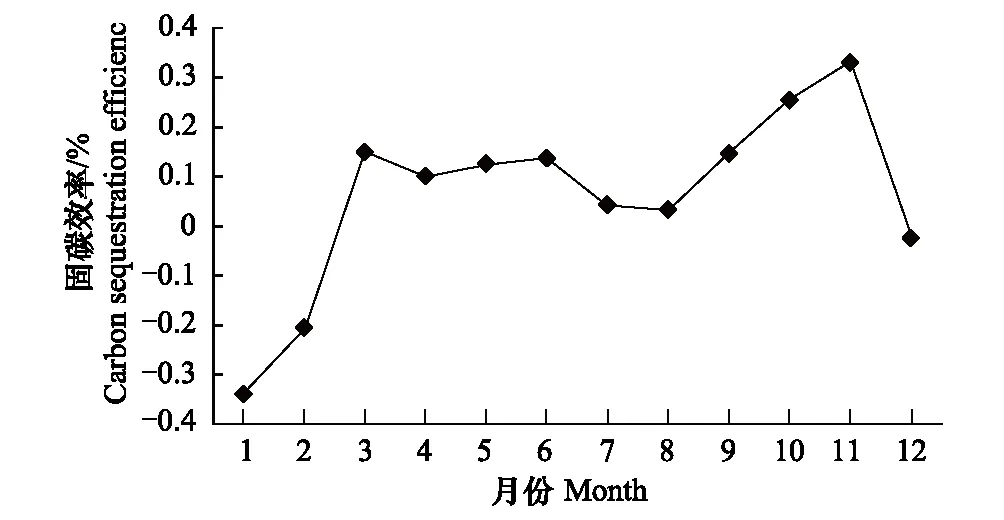
图3 固碳效率变化
2.3 竹林固碳效率
在一定的时间尺度上,生态系统碳交换通量与碳固定和排放速率根据选定的研究界面和对象能够相互直接转化[24],在数值上GEE可等同为生态系统固碳速率,RE即为生态系统碳排放速率,NEE即生态系统净固碳速率,量纲不变仍为gC·m-2·a-1。相对于植物生理研究,定义有碳利用效率(CUE),就是植物净积累的碳占总光合生产中固定碳的比例[25],即有CUE=NPP/GEP[26],该指标表示植物碳同化能力的强弱,但是对于整个生态系统的碳收支研究缺失土壤呼吸和植物凋落物等异养呼吸碳排放计算,不能代表整个生态系统碳收支水平。如果将大气-植被-土壤看作一个连续的整体,研究在生态系统气体交换过程中生态系统的碳收支,将生态系统净固碳速率与生态系统固碳速率比值,可以暂时定义为生态系统碳固定效率(ECSE),反映生态系统气体交换中固定下来碳的比例,即ECSE=NEE/GEE。从图3中看出,生态系统的碳利用效率从1月份开始为负值表明,是极强的碳释放,冬季雷竹虽然有一定的光合能力但生态系统呼吸大大超过光合生产的净积累量;3月开始,变为正值转,为固碳并保持在10—16%;夏季7—8月固碳能力下降,碳排放的比例增加,进入秋季达到峰值,有生态系统气体交换过程有33%被固定下来,全年固碳效率为11%。
2.4 高效经营覆盖措施对CO2通量的影响

图4 土壤5cm温度与空气1m温度的比较
冬季覆盖技术是雷竹高效经营的关键技术,覆盖的主要作用是增加土壤温度。夏季土壤表层温度高于深层温度,在冬季则相反,土壤温度随深度增加而升高。在冬季增加覆盖措施后5cm土壤温度要高于1m处气温5—10℃。从图4中看出在12—2月,地表5cm土壤温度明显高出空气温度,平均温度都维持在10℃以上,这就加大了生态系统的呼吸,同时冬季光合有效辐射为全年最低,形成冬季月份碳源。
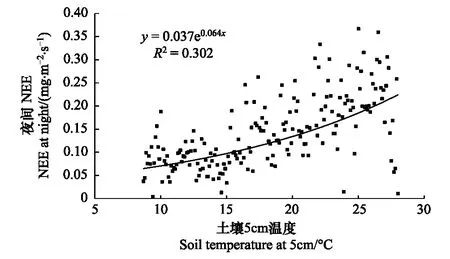
图5 土壤5cm温度变化与夜间NEE的关系
全年土壤温度变化范围9—26℃,波动范围不大,夜间NEE可以代表生态系统呼吸即包括植物暗呼吸、土壤微生物呼吸以及凋落物分解的CO2排放,最高呼吸在0.6—0.7mg·m-2·s-1。全年的土壤5cm温度与经摩擦风速筛选后夜间NEE关系并不明显,因为点数太多,将各个温度下的夜间NEE做平均处理,得到图5。从图5中,可以看出生态系统与土壤5cm成指数关系,但相关系数只有0.3,成指数关系这与之前有关文献报道的研究结果较一致[16,27]。ChinaFLUX站点也有相似的报道,陈述悦研究华北麦田发现5cm地温同土壤呼吸相关性最好[28],刘允芬等研究千烟洲人工针叶林土壤温度与夜间NEE数值呈指数关系相关系数0.38[29]。相关系数不大的原因可能有两个,首先冬季的覆盖增温措施干扰了土壤自然的呼吸,使温度变化范围减小,生物菌肥的使用加大了呼吸;再有温度升高使植物自养呼吸增加,生态系统呼吸也随之增加。
假定雷竹光合能力不变,数据显示冬季无覆盖时近地1m空气温度与土壤5cm温度相差不大,用1m处空气温度代替无覆盖的5cm土壤温度,根据与夜间NEE的指数方程模拟无覆盖的生态系统呼吸。以温度为影响因子比较无覆盖与有覆盖的碳排放,模拟结果说明12—2月单位面积上,覆盖后分别多增加21.56g,26.52g,20.53g的排放(表3)。因此若雷竹林无覆盖可少排放CO2,相应冬季月份可由碳源转汇。
冬季覆盖增加的碳排放部分来自覆盖的有机物分解。从表4中可以计算不包括地下部分生长计算碳收支平衡,其中雷竹每株干重计以3kg计算,每年每亩伐300株,地上部分碳密度参照周国模研究毛竹碳密度[19]以500g/kg计算,竹笋收获以1500kg,含水90%计算,总年收获量为525kg碳,即787.50Cg·m-2·a-1。竹笋收获占14%,竹材收获占86%,竹材的碳可以作为碳库多年储存,竹笋的碳运出系统后又将排放到大气中,则雷竹林系统为十分微弱的碳源。雷竹林NEE较水稻田、北方农田都小,但收获量缺明显高出其他农田系统,可见高效经营措施对收获量产出的影响。
覆盖物投入的有机碳含量以及砻糠收回,稻草则腐烂入土的比例需要进一步实验,对竹林系统碳库影响都还不甚清楚。目前有关覆盖物对雷竹林碳储量影响、雷竹土壤有机质转化研究的文献还很少见,初步估计,每年每公顷投入覆盖的稻草及砻糠等有机物料共100 t左右,大量的有机质输入对土壤有机碳库的组成分解及转化应产生重要影响[30-32]。有文献研究15a种植雷竹有机碳转化变化,覆盖经营后极大地提高了土壤有机质含量,每年每亩表层有机质增加达到616 kg,占投入物料的23.8%。该值略低于南方红壤区作物秸秆的平均腐殖化系数(0.22—0.34,平均 0.28),可能是由于砻糠 C/N 比较高,不易腐殖化[33]。每亩1.6t有机碳投入,以23.8%转化系数计算,将有380kg有机碳进入系统,即增加了碳排放571Cg·m-2·a-1,这些有机碳在系统中如何分配的有待深入研究。

表3 模拟覆盖与无覆盖碳排放变化

表4 高效经营中投入产出(以666.7m2计)
3 结论与展望
本文分析了太湖源人工经营的雷竹林碳通量各月的平均日变化趋势,季节碳通量的变化趋势,同时探究了冬季覆盖增温过程中对碳通量变化以及碳平衡的影响。研究表明:
人工经营的雷竹林生态系统全年各月碳通量平均日变化趋势明显;2010年10月到2011年9月碳收支为-126.303gC·m-2·a-1,RE为1108.845 gC·m-2·a-1,GEE为-1235.15gC·m-2·a-1。整体为碳汇,冬季覆盖的12—2月份为碳源,其余月份为碳汇。通量过程具有明显的季节性,雷竹林生态系统月尺度上的NEE符号变化时刻和最大的碳排放、碳吸收时刻都有明显的差异,通量符号由正值转为负值在日出1h—1.5h,春冬季与夏秋季相比延迟1h;由负值转为正值时刻较集中,发生在17:00—18:00;最大碳汇时刻秋季在11:00,冬季在12:00,春夏介于11:00—12:00之间。温度是影响碳通量的主要因子,5cm处土壤温度与生态系统呼吸存在明显的指数关系。在春季NEE随NEE与RE的同时升高而增加,在秋季NEE随NEE与RE的同时下降而增加,出现两个峰值并有碳汇总量秋季大于春季。秋季竹林NEE贡献达68.9%,同时固碳效率最高为33%。冬季覆盖通过投入有机物料,提高地温,同时增大了生态系统呼吸,增加了碳排放。
竹林具有很强的森林碳汇潜力,合理的耕作措施,科学采伐老竹,保持生态系统活力,可为陆地生态系统维持碳氧平衡应对气候变化做出积极的贡献。另一方面,对于竹林生态系统碳循环研究,本文仅对雷竹林的碳源汇做了结论,对与影响碳吸收、排放的各个分量未做计算。同时本站点仅对雷竹林一年的碳通量过程研究进行初步观测研究,时间较短,因碳通量过程有明显的年际变化易受气候变化影响,要通过长期的连续观测和相应的辅助实验,来做深入研究。
[1]Tans P P,Fung I Y,Takahashi T.Observation constraints on the global atmospheric CO2budget.Science,1990,247(4949):1431-1438.
[2]Berbigier P,Bonnefond J M,Mellmann P.CO2and water vapour fluxes for 2 years above Euroflux forest site.Agricultural and Forest Meteorology,2001,108(3):183-197.
[3]Yu G R,SunX M.Flux Measurement and Research of Terrestrial Ecosystem in China.Beijing:Science Press,2008.
[4]Yu G R,Sun X M.Principles of Flux Measurement in Terrestrial Ecosystems.Beijing:Higher Education Press,2006:4-4.
[5]Payeur-Poirier J L,Coursolle C,Hank A,Giasson M A.CO2fluxes of a boreal black spruce chronosequence in eastern North America.Agricultural and Forest Meteorology,2012,153:94-105
[6]FangW,He J C,Lu K X,Chen J H.Cultivation techniques of early shooting and high yielding for Lei Bamboo Sprout.Journalof Zhejiang Forestry College,1994,11(2):121-128.
[7]Wang Z T,FangW,He J C.High Yield Cultivation Techniques of Lei Bamboo shoots.Beijing:China Forestry Press,1993.
[8]Xu L L,Hu Y B,Hu Y C,Sun C M,Yu M C,Liang K,Lang J B.Lei bamboo production development and covered technology.Shanghai Agricultural Science and Technology,2008,(2):85-86.
[9]Baldocchi D D,Hicks B B,Meyers T P.Measuring biosphere-atmosphere exchanges of biologically related gases with micrometeorological methods.Ecology,1988,69(5):1331-1340.
[10]McMillen R T.An eddy correlation technique with extended applicability to non-simple terrain.Boundary-Layer Meteorology,1988,43(3):231-245.
[11]FokenTh,Wichura B.Tools for quality assessment of surface-based flux measurements.Agricultural and Forest Meteorology,1995,78(1/2):83-105.
[12]Paw U K T,Baldocchic D D,Meyers T P,Wilson K B.Correction of eddy-covariance measurements incorporating both advective effects and density fluxes.Boundary-Layer Meteorology,2000,97(3):487-511.
[13]Reynolds O.On the dynamical theory of incompressible viscous fluids and the determination of the criterion.Philosophical Transactions of the Royal Society of LondonA,1895,186:123-164.
[14]Garratt J R.Limitations of the eddy-correlation technique for the determination of turbulent fluxes near the surface.Boundary-Layer Meteorology,1975,8(3/4):255-259.
[15]Song X,Liu Y F,Xu XF,Yu GR,Wen X F.Comparison study on carbon dioxide,water and heat fluxes of the forest ecosystem in red earth hilly zone over winter and spring.Resources Science,2004,26(3):96-104.
[16]FalgeE,Baldocchi D,Olson R,Anthoni P,Aubinet M,Bernhofer C,Burba G,Ceulemans R,Clement R,Dolman H,Granier A,Gross P,Grünwald T,Hollinger D,Jensen N O,Katul G,Keronen P,Kowalski A,Ta Lai C,Law B E,Meyers T,Moncrieff J,Moors E,Munger J W,Pilegaard K,Rannik Ü,Rebmann C,Suyker A,Tenhunen J,Tu K,Verma S,Vesala T,Wilson K,Wofs S.Gap filling strategies for defensible annual sums of net ecosystem exchange.Agricultural and Forest Meteorology,2001,107:43-69.
[17]Katul G,Lai C T,Schäfer K,Vidakovic B,Ellsworth D,Oren R.Multiscale analysis of vegetation surfacefluxes:from seconds to years.Advances in Water Resources,2001,24(9/10):1119-1132.
[18]Wilson K B,Baldocchi D D.Comparing independent estimates of carbon dioxide exchange over 5 years at a deciduous forest in the southeastern United States.Journal of Geophysical Research,2001,106(D24):34167-34178.
[19]Zhou G M.Carbon Storage,Fixation and Distribution in Mao Bamboo(Phyllostachyspubescens) Stands Ecosystem.Zhejiang:Zhejiang University,2006.
[20]Li J J,Pan G X,Li LQ,Zhang X H.Estimation of net carbon balance and benefits of rice-rice cropping farm of a red earth paddy under long term fertilization experiment from Jiangxi,China.Journal of Agro-Environment Science,2009,28(12):2520-2525.
[21]LiJ,YuQ,Sun X M,Tong X J,Ren C Y,Wang J,Liu E M,Zhu Z L,Lin G R.The north China plain farmland ecological system and its environmental regulation mechanism carbon exchange.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):210-223.
[22]Liu Y F,Song X X,Sun X M,Wen X F,Chen Y R.CO2flux seasonal changes and environmental factors of influence form Qianyanzhou of Coniferous.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):109-117.
[23]Zhou G M,Jiang P K,Xu Q F.Bamboo Forest Ecosystem Carbon Fixation and Transformation.Beijing:Science Press,2010.
[24]Yu G R,Wang Q F,Liu Y C,Liu Y H.Conceptual framework of carbon sequestration rate and potential increment of carbon sink of regional terrestrial ecosystem and scientific basis for quantitative carbon Authentification.Progress in Geography,2011,30(7):772-774.
[25]Cabral O M R,Gash J H C,Rocha H R,Marsden C,Ligo M A V,Freitas H C,Tatsch J D,Gomes E.Fluxes of CO2above a plantation of Eucalyptus in southeast Brazil.Agricultural and Forest Meteorology,2011,151(1):49-59.
[26]ChoudhuryB J.Modeling radiation-and carbon-use efficiencies of maize,sorghum,and rice.Agricultural and Forest Meteorology,2001,106(4):317-330.
[24]LloydJ,Taylor J A.On the temperature dependence of soil respiration.Functional Ecology,1994,8(3):315-323.
[27]LloydJ,Taylor J A.On the temperature dependence of soil respiration.Functional Ecology,1994,8(3):315-323.
[28]ChenSS,LiJ,Lu P L,Wang Y H,Yu Q.Soil respiration characteristics in winter wheat field in North China Plain.Chinese Journal of Applied Ecology,2004,15(9):1552-1560.
[29]Liu Y F,Yu G R,Wen X F,Wang Y H,Song X,Li J,Sun X M,Yang F T,Chen Y R,Liu Q L.The season of CO2flux variation characteristics form Subtropical plantation ecological system.Science in ChinaSeries D:Earth Sciences,2006,36(Supplement):91-102.
[30]Jiang P K,Zhou GM,Xu Q F.Effect of intensive cultivation on the carbon pool of soil inPhyllostachyspraecoxstands.Scientia Silvae Sinicae,2002,(6):6-11
[31]Jiang P K,Xu Q F.Effects of fertilization on soil active organic carbon underPhyllostachyspraecoxstand.Chinese Journal of Applied Ecology,2005,(2):253-256.
[32]Cai R R,HuangF,Sun D,Qin H,Yang F,Zhuang S Y,Zhou G M,Cao Z H.Temporal and spatial variation of soil organic matters inPhyllostachyspraecoxstands with intensive cultivation management.Journal of Zhejiang Forestry College,2007,(4):450-455.
[33]Li G D,Liu G Q,Zhuang S R,Gui R Y.Changes of organic matter in soils planted Lei Bamboo with different years.Chinese Journal of Soil Science,2010,41(4):847-848.
参考文献:
[3]于贵瑞,孙晓敏,等.中国陆地生态系统碳通量观测技术及时空变化特征.北京:中国科学出版社,2008.
[4]于贵瑞,孙晓敏.陆地生态系统通量观测的原理与方法.北京:高等教育出版社,2006:4.
[6]方伟,何钧潮,卢可学,陈健华.雷竹早产高效栽培技术.浙江林学院学报,1994,11(2):121-128.
[7]汪祖潭,方伟,何钧潮.雷竹笋用林高产栽培技术.北京:中国林业出版社,1993.
[8]徐缪畅,胡永斌,胡余楚,孙春明,俞茂昌,梁侃,朗进宝.雷竹生产发展与竹园覆盖技术.上海农业科技,2008,(2):85-86.
[15]宋霞,刘允芬,徐小锋,于贵瑞,温学发.红壤丘陵区人工林冬春时段碳、水、热通量的观测与分析.资源科学,2004,26(3):96-104.
[19]周国模.毛竹林生态系统中碳储量、固定及其分配与分布的研究[D].浙江:浙江大学,2006.
[20]李洁静,潘根兴,李恋卿,张旭辉.红壤丘陵双季稻稻田农田生态系统不同施肥下碳汇效应及收益评估.农业环境科学学报,2009,28(12):2520-2525.
[21]李俊,于强,孙晓敏,同小娟,任传友,王靖,刘恩民,朱治林,于贵瑞.华北平原农田生态系统碳交换及其环境调控机制.中国科学D辑:地球科学,2006,36(增刊):210-223.
[22]刘允芬,宋霞,孙晓敏,温学法,陈永润.千烟洲人工针叶林CO2通量季节变化及其环境因子的影响.中国科学D辑:地球科学,2004,34(增刊):109-117.
[23]周国模,姜培坤,徐秋芳.竹林生态系统中碳的固定与转化.北京:科学出版社,2010.
[24]于贵瑞,王秋凤,刘迎春,刘颖慧.区域尺度陆地生态系统固碳速率和增汇潜力概念框架及其定量认证科学基础.地理科学进展,2011,30(7):772-774.
[28]陈述说,李俊,陆佩玲,王迎红,于强.华北平原麦田土壤呼吸特征.应用生态学报,2004,15(9):1552-1560.
[29]刘允芬,于贵瑞,温学法,王迎红,宋霞,李菊,孙晓敏,杨风亭,陈永瑞,刘琪瓃.千烟洲中亚热带人工林生态系统CO2通量的季节变异特征.中国科学D辑:地球科学,2006,36(增刊):91-102.
[30]姜培坤,周国模,徐秋芳.雷竹高效栽培措施对土壤碳库的影响.林业科学,2002,(6):6-11.
[31]姜培坤,徐秋芳.施肥对雷竹林土壤活性有机碳的影响.应用生态学报,2005,(2):253-256.
[32]蔡荣荣,黄芳,孙达,秦华,杨芳,庄舜尧,周国模,曹志洪.集约经营雷竹林土壤有机质的时空变化.浙江林学院学报,2007,(4):450-455.
[33]李国栋,刘国群,庄舜尧,桂仁意.不同种植年限下雷竹林土壤的有机质转化.土壤通报,2010,41(4):847-848.
