栎属7种植物种子的发芽抑制物质研究
2013-12-21李庆梅刘广全侯龙鱼胡金鑫
李庆梅,刘 艳,,刘广全 ,刘 勇,侯龙鱼,胡金鑫
(1.林木遗传育种国家重点实验室,中国林业科学研究院林业研究所,北京 100091;2.北京林业大学森林培育与保护教育部重点试验室,北京 100083;3.中国水利水电科学研究院,北京 100048;4.西北农林科技大学,杨凌 712100;5.中国科学院植物研究所植被与环境变化重点实验室,北京 100093)
栎属(Quercus)是我国重要的硬阔叶树种,具有耐寒、耐旱和耐瘠薄的生态特性[1]。麻栎(Q.acutissima Carruth.)、锐齿栎(Q.aliena var.acuteserrata Maxim ex Wenz.)、栓皮栎(Q.variabilis BI.)和蒙古栎(Q.mongolica Fischer ex ledebour.)等,在土壤改良,植被恢复、水土保持、水源涵养等方面具有重要作用[2]。近年来,随着经济的发展,栎属植物已成为主要的经济和能源树种,生物量和蓄积量在林木中占较大的比重[3]。
栎类植物人工造林生产实践中通常采用播种育苗,但存在延迟萌发、出苗不整齐的现象[4]。自然条件下,最先出苗和后出苗的相差可达数周[5],出苗整齐性很差[6]。近几年栎类植物经常采用容器育苗[7],这就要求苗木快速整齐的萌发,这也是提高育苗质量的一个有效途径[8]。因此,研究和认识造成栎类种子延迟萌发和出苗不整齐的原因并找出相应的改善途径,不仅可提高育苗的质量,改善栎林自然更新率低的现状,对育苗具有一定的指导意义。
栎类种子延迟萌发和出苗不整齐的原因主要包括:(1)形态影响即种子大小和种皮机械束缚影响[4,9];(2)次生代谢物如单宁酸和酚类等通过抑制一些代谢途径影响种子的萌发[10];(3)植物激素脱落酸(ABA)和植物生长调节剂(IAA)影响种子的代谢途径[11-12]。发芽抑制物质是由植物体产生的,或是植物体中虽不存在而与发芽抑制有关的物质,这类物质可延迟或抑制同种或异种种子的发芽[13]。Peterson[6]发现成熟水栎果皮的水提物中含有抑制物质。为了抵御昆虫取食,栎类种子能产生抑制种子萌发的次生代谢物,如多酚(polyphenols)[10]和单宁酸(tannins)[14]。但对将栎类种子解剖成不同部位,并采用科学有效的方法提取其抑制物质的研究较少。系统溶剂法和生物测定法是测定和分析种子中抑制物质的有效方法[15-17],且该方法在栎类种子抑制物质研究上应用较少,本研究通过对7种栎属植物种子萌发抑制物质的研究,试图找出导致栎属种子延迟萌发和出苗不整齐的原因,为生产上采取措施提高种子发芽率和整齐性提供理论依据。
1 材料与方法
1.1 试验材料
试验用种子来源于中国科学院北京植物园、河南省小秦岭自然保护区和陕西省秦岭东段新开岭自然保护区,各地区所采物种和气象因子见表1。
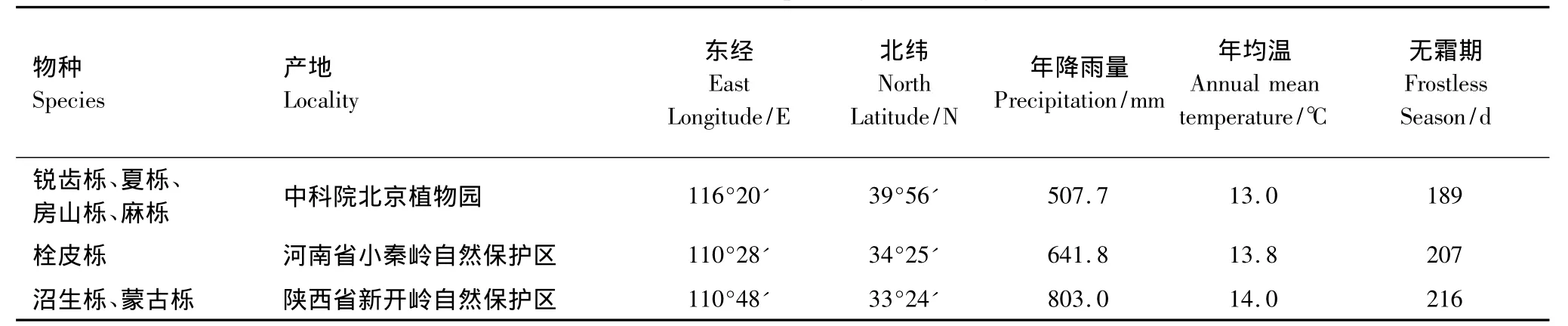
表1 采种地点及相应气象因子Table 1 Seed sites and corresponding meteorological factors
生物活性测定所用白菜(Brassica pekinensis)种子品种为中白81,由北京中蔬园艺良种研究开发中心提供,纯度96%,净度98%,发芽率85%以上,含水量约7%。采收于2010年6月,保存在(3±1)℃的冰箱中。
1.2 研究方法
1.2.1 种子预处理和基本形态指标测定
采收后的种子用自来水浸泡,5 min后淘汰仍漂浮在水面的种子[18],剩下的种子(将壳斗即果碗和种子分开)擦拭干表面水分后平铺自然阴干,间隔2—3 h翻动1次,2 d后将种子置于(3±1)℃贮藏柜中保存。
1.2.2 种子发芽试验
为了验证栎属植物种子延迟萌发的现象,随机选取麻栎、夏栎和房山栎各45粒,15粒1组,共3个重复。将种子放入铺有3层滤纸的培养皿中[4,19]置于25℃、光照8 h培养箱进行发芽试验。种子萌发后每7 d统计1次,分别统计出根数和出芽数,并计算出根率和发芽率,计算公式如下:
出根率(%)=出根种子数/供试种子数×100%
发芽率(%)=发芽种子数/供试种子数×100%
1.2.3 栎属7种植物种子不同部位甲醇浸提液的提取及生物活性测定
将7种种子解剖为种皮(胚和子叶外围的部分)、种胚(胚及周围部分子叶)和子叶(远离胚端2/3子叶)3个部分,30℃烘干后用粉碎机磨碎,过1 mm筛。分别称取50 g加入足量的80%甲醇溶液,于(3±1)℃条件下密闭浸提,并间隔4 h搅拌1次,使其充分浸提[16]。每24 h更换1次浸提液,重复浸提3次。合并浸提液,离心,取上清液,减压浓缩至1 g/mL。分别取一定体积浓缩液稀释至0.5 g/mL,每个浓度10 mL,贮于(3±1)℃条件下备用。
在直径9 cm的培养皿中铺入滤纸,分别加入3 mL待测液,待各相有机溶剂完全挥发(水相除外)后,每皿内再加入3 mL蒸馏水,并以同体积蒸馏水作为对照。白菜种子用始温45℃水浸泡10 min后,取100粒均匀摆放在培养皿中,置于25℃恒温光照培养箱内,48 h统计白菜种子的发芽率(胚根长度大于白菜种子直径时即为发芽),72 h测其苗高和根长。每个处理3个重复。

图1 种皮、子叶和胚中抑制物质提取流程图Fig.1 The flow chart of inhibitor extract in pericarp,cotyledon and embryo
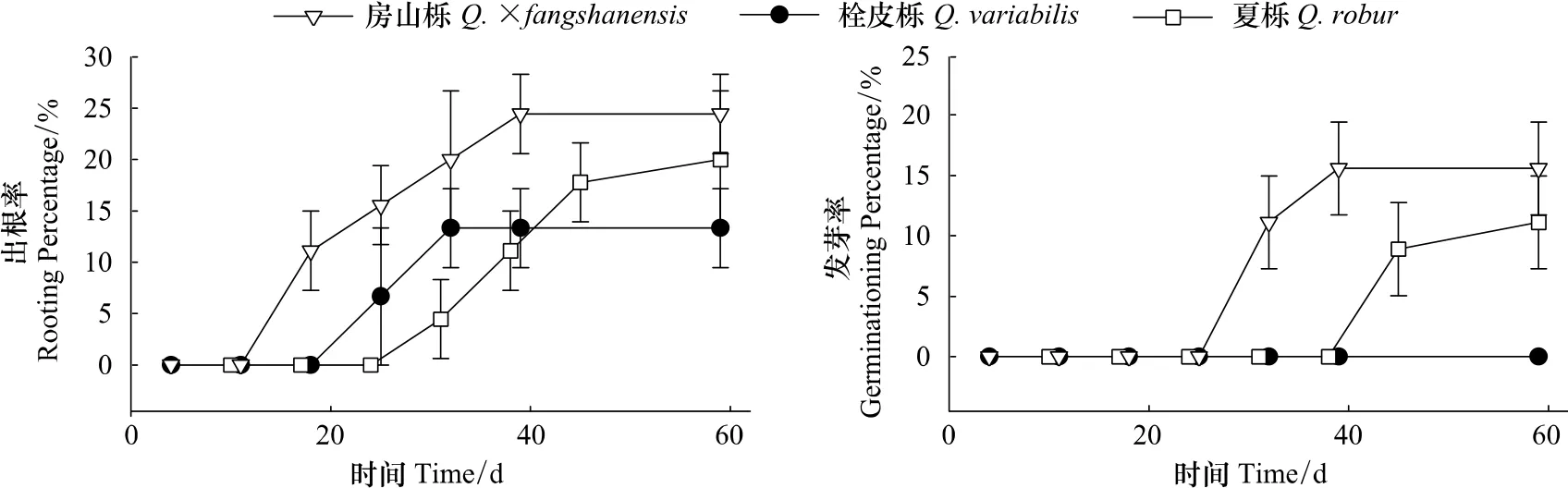
图2 麻栎、夏栎和房山栎种子萌发过程Fig.2 The process of germinating of Q.acutissima,Q.robur and Q.×fangshanensis seeds
1.2.4 栎属7种植物种种子不同部位中抑制物质的初步分离及生物活性测定
按系统溶剂法[15],将种皮、子叶和种胚的甲醇浸提液分离为石油醚相、乙醚相、乙酸乙酯相、甲醇相和水相(图3)。然后将各有机相溶液分别置于旋转蒸发仪RE-52AA上进行浓缩蒸发,最后将各有机相定容至1 g/mL,置于(3±1)℃冰箱中保存备用。各分离相生物活性测定方法同1.2.3。

图3 不同浓度种子甲醇浸提液对白菜种子发芽率的影响Fig.3 Effects of different concentrations of methanol extract from embryo,cotyledon and pericarp of seeds on the germination percentage of B.pekinensis seeds
1.3 数据分析
用SPSS11.5进行单因素方差分析,种子发芽率方差分析前经反正弦转换,用Schefffe检验其差异显著性。结果以平均值±标准差(Mean value±SD)表示。图由SigmaPlot 10.0辅助完成。
2 结果与分析
2.1 栎属3种种子萌发过程
麻栎、夏栎(Q.robur L)和房山栎(Q.×fangshanensis Liou)从最初出根到出根末期分别用了14、28和28 d,最后的出根率分别为(13.3±3.8)%、(20.0±6.7)%和(24.4±3.8)%。夏栎和房山栎种子开始发芽的天数比出根的时间大约推迟了14 d,且发芽率均小于出根率,分别为(11.1±3.8)%、(15.6±3.8)%。麻栎种子在培养期内没有发芽(图2)。3个物种的生活力(TTC染色法测定)均超过80%,但出根率和发芽率均不超过30%,说明3种栎属种子存在延迟萌发、只生根不萌芽、出苗不整齐等现象。
2.2 不同浓度的种子甲醇浸提液对白菜种子发芽的影响
7种栎属种子的种皮、种胚和子叶甲醇浸提液对白菜种子发芽率表现出不同影响(图3)。栓皮栎、锐齿栎、蒙古栎、沼生栎(Q.palustris Muench)和麻栎种胚浸提液1.0 g/mL浓度显著降低了白菜种子发芽率,比对照发芽率分别降低了83%、26%、26.3%、18.3%和17%;夏栎和房山栎分别降低了9.33%和10%,与对照没有显著差异。只有栓皮栎的0.5 g/mL浓度的种胚浸提液显著降低了白菜种子的发芽率,其它物种虽有不同程度的降低,但与对照差异不显著(图3)。
对于子叶甲醇浸提液,1.0 g/mL浓度时栓皮栎、锐齿栎和蒙古栎以及0.5 g/mL浓度时栓皮栎显著降低了白菜种子发芽率,其它均没有显著影响(图3)。
种皮的甲醇浸提液,只有栓皮栎和蒙古栎1.0 g/mL浓度显著降低了白菜种子发芽率(图3)。
因此,7个物种中,栓皮栎各部分甲醇浸提液抑制作用最强,其次是锐齿栎和蒙古栎,夏栎和房山栎的抑制作用最弱。种子不同部位,抑制作用表现为胚>子叶>种皮,且浓度提高抑制作用增强。
2.3 不同浓度种子甲醇浸提液对白菜种子萌发后形成的幼苗苗高和根长的影响

图4 不同浓度的供试种子不同部位甲醇浸提液对白菜种子苗高和根长的影响Fig.4 Effects of different concentrations of methanol extract from embryo,cotyledonand pericarp of Quercus seeds on the shoot and root length of B.pekinensis seeds
由图4可以看出,与对照相比,1.0 g/mL和0.5 g/mL浓度7树种种胚甲醇浸提液均显著降低了白菜种子的苗高和根长,且高浓度的抑制作用大于低浓度。栓皮栎、锐齿栎、蒙古栎和沼生栎1.0 g/mL浓度种胚甲醇浸提液对白菜种子苗高的抑制作用显著高于麻栎、夏栎和房山栎;栓皮栎和锐齿栎0.5 g/mL浓度种胚甲醇浸提液白菜种子苗高显著低于其它树种,抑制作用较强;各树种间2种浓度种胚甲醇浸提液对白菜种子根长没有显著性差异,栓皮栎和锐齿栎的相对较小。
与对照相比,7个树种的子叶甲醇浸提液均降低了白菜种子的苗高和根长。栓皮栎和锐齿栎子叶甲醇浸提液白菜种子的苗高最低,说明它们对苗高的抑制作用最强,蒙古栎对白菜种子苗高的抑制作用最弱,其他树种的抑制作用无显著差异。各树种间对根长的抑制作用差不多。高浓度的苗高和根长比低浓度的低,具有较强的抑制作用(图4)。
图4结果表明,锐齿栎种皮两个设定浓度的甲醇浸提液均显著降低白菜种子的苗高,栓皮栎、麻栎、夏栎、沼生栎和蒙古栎种皮两个设定浓度的甲醇浸提液一定程度上降低了白菜种子苗高,且高浓度的抑制作用强于低浓度;房山栎2种浓度白菜种子的苗高与对照差不多。栓皮栎和锐齿栎2种浓度显著降低了白菜种子的根长,其它树种的甲醇浸提液也一定程度抑制了白菜种子根长的生长。
7个树种的种胚、子叶和种皮的甲醇浸提液对苗高的影响差异不大,胚和子叶的甲醇浸提液对根长的抑制作用大于种皮。
2.3 各有机相对白菜种子萌发的影响
各树种甲醇相对白菜种子的发芽率抑制作用较强(图5),栓皮栎、锐齿栎和夏栎种胚以及蒙古栎子叶的的甲醇相均显著降低了白菜种子的发芽率,分别降低了25.0%、17.3%、18.3%和15.3%。其它4个有机相,只有沼生栎种皮的乙醚相对白菜种子发芽率有显著抑制作用。
各树种甲醇相对白菜种子苗高和根长也有显著抑制作用,种皮对苗高没有显著影响。只有栓皮栎胚的甲醇相对苗高的抑制作用明显高于子叶,其余树种胚和子叶甲醇相对苗高和根长的影响差异不显著。
锐齿栎种胚的乙酸乙酯相对苗高有显著抑制作用,其他树种不同部位没有显著抑制白菜种子苗高的生长。锐齿栎、麻栎和夏栎种子3个部位以及栓皮栎、蒙古栎和房山栎种胚和子叶的乙酸乙酯相均不同程度降低了白菜种子的根长,但与对照差异不显著,因此,种胚与子叶各有机相对白菜种子根长的抑制作用大于种皮。各树种的石油醚相、乙醚相和水相对白菜种子的苗高和根长基本没有影响。
综合发芽率、苗高和根长指标看,各树种甲醇相的抑制作用最强,其次是乙酸乙酯相,而且种胚和子叶的抑制作用较种皮强,其他相的抑制作用不是很明显。
3 结论与讨论
栓皮栎、锐齿栎、蒙古栎、沼生栎和麻栎甲醇浸提液均能显著降低白菜种子发芽率,且抑制作用逐渐降低,说明它们的甲醇浸提液均含有萌发抑制物质。Peterson[6]在水栎(Quercus nigra)果皮的水提物中也发现含有抑制物质,燕山红栗(Castanea mollissima B I cv.‘Yanshanhong’)和板栗(C.mollissima Blume)种皮中也含有抑制物质[20-21]。有研究认为为了抵御昆虫取食,栎类种子产生次生代谢物[9],如多酚(polyphenols)[10]和单宁酸(tannins)[14],这些物质能抑制种子的萌发。对多种栎类的统计表明休眠的红栎类(Red oaks)种子单宁酸含量明显多于无休眠的白栎类(White oaks)种子[22]。
夏栎和房山栎甲醇浸提液对白菜种子发芽率没有显著影响,但对根长和苗高有一定的抑制作用。Bonner and Vozzo[4]研究指出白栎亚属的大部分种子没有休眠,但在萌发过程中,出苗不整齐。由此可见,栎类种子的延迟萌发和出苗不整齐受多种物质的影响,在物种间存在着差异。

图5 种子各部位各有机相浸提液对白菜种子萌发的影响Fig.5 Effects of inhibitors of pericarp,embryo and cotyledon from 5 phases on the germination percentage,root length and shoot height of B.pekinensis
栎属7个种的种子不同部位甲醇相均能显著降低白菜种子发芽率、苗高或根长,说明甲醇相的抑制作用最强,其次是乙酸乙酯相,其他相的抑制作用不是很明显,由此推断栎属种子中所含抑制物种可能为极性化合物。其它林木物种种子不同部位,如西洋参(Panax quinque folium L.)果肉[15]、珙桐(Davidia involucrata Baill.)果皮和胚乳[23]、南京椴(Tilia miquelian Maxim.)果皮和种子[24]、南方红豆杉(Taxus ch inensis var.m airei ChengetL.K.Fu)种皮和胚乳[16]、山茱萸(Macrocarpiumofficinale Nakai)外果皮和种壳[25]和美国桂花(Osmanthus americanus)种壳和胚乳[26]的甲醇相均对白菜种子的发芽率或生长有显著抑制作用。其中,珙桐果皮[22]、南方红豆杉种皮和胚乳[16]和山茱萸外果皮和种壳[25]的乙酸乙酯相都含有抑制物质。另外,西洋参果肉[15]、珙桐果皮和胚乳[23]、南京椴果皮[24]、南方红豆杉种皮和胚乳[16]、美国桂花种壳和胚乳的乙醚相[26]以及南方红豆杉种皮和胚乳的石油醚相和水相[16]也均含有抑制物质。甲醇相中均含有抑制物质可能是上述各树种种子在贮藏和其萌发过程中的主要生理过程均相似,为了适应环境和完成生长发育,自身会产生一些化合物,抑制周围其它种子萌发或自身萌发。种子适应不同环境条件可能是造成抑制物质有差别的原因,如有些休眠是为了错开寒冷的环境,有些是为了预防干旱和驱避虫食。种子中可能存在多种抑制物,但起关键作用的可能仅有一种或几种[23]。本试验7种栎属植物种子抑制物测定均以白菜种子的萌发和生长为参照,抑制物对栎类种子自身萌发和生长的抑制程度以及抑制机理有待进一步研究。
试验证明栎类种子中含有抑制物质,这些抑制物质主要抑制种子萌芽和生长。研究发现,栎类植物存在延迟萌发、出苗不整齐[4-5]和只生根不萌芽现象[27],麻栎、夏栎和房山栎种子的发芽试验进一步验证了这种现象。除了种皮的机械束缚[9]和子叶的影响外[7],栎类种子中含有抑制物质也可能是造成其延迟萌发和出苗不整齐的原因。进一步鉴定抑制物质及其抑制机制,研究减弱和消除其对萌发的抑制、促进种子的快速萌发和整齐出苗的途径,对栎属植物造林生产和栎类更新均具有重要意义。
[1]Li W Y,Wang B,Li G C.Ecology benefits,economical values and conservation strategies of Quercus species.Forest Science and Technology,2001,(8):13-15.
[2]Zhang H M,Wang X J,Yan JH,Bai JH,Lu Y G.Analysis on spatial distribution and growth process of oak forests in Beijing.Journal of Beijing Forestry University,2010,32(Supplement):71-79.
[3]Gao J R.Hydrological effect of Quercus aliena var.acuteserrata stands in forest region of the Qinling Mountains.Journal of Beijing Forestry University,1998,20(6):13-15.
[4]Bonner F T,Vozzo J A.Seed biology and technology of Quercus.New Orleans:USDA Forest Service,1987.
[5]Suszka B,Muller C,Bonnet-Masimbert M.Nasiona Drzew Les'nych:Od zbioru do siewu.[Seeds of forest broadleaves from harvest to sowing].PWN:Warszawa-Poznań,2000:307.
[6]Peterson J K.Mechanisms involved in delayed germination of Quercus nigra L.seeds.Annals of Botany,1983,52(1):81-92.
[7]Giertych M J,Suszka J.Consequences of cutting off distal ends of cotyledons of Quercus robur acorns before sowing.Annals of Forest Science,2011,68(2):433-442.
[8]Suszka B.Generative propagation//Bugała W,ed.Dęby(Quercus robur L;Q petraea(Matt)Liebl)-Nasze drzewa les'ne.Poznań:Bogucki Wydawnictwo Naukowe,2006:305-388.
[9]Quero J L,Villar R,Marañón T,Zamora R,Poorter L.Seed-mass effects in four mediterranean Quercus species(Fagaceae)growing in contrasting light environments.American Journal of Botany,2007,94(11):1795-1803.
[10]Rakic'S,Povrenovic'D,Teševic'V,Simic'M,Maletic'R.Oak acorn,polyphenols and antioxidant activity in functional food.Journal of Food Engineering,2006,74(3):416-423.
[11]Finch-Savage W E,Clay H A.Water relations of germination in the recalcitrant seeds of Quercus robur L.Seed Science Research,1994,4(3):315-322.
[12]Suszka B,Muller C,Bonnet-Masimbert M.Seeds of forest broadleaves:from harvest to sowing.Paris:Institute National de la Recherche Agronomique(INRA),1996.
[13]Evenari M.Germination inhibitors.The Botanical Review,1949,15(3):153-194.
[14]Shimada T.Nutrient compositions of acorns and horse chestnuts in relation to seed-hoarding.Ecological Research,2001,16(4):803-808.
[15]Huang Y G,Cui SY,Lu Q,Niu H B,Zhao CY,Han B R,Yang JX,Li X G.Growth inhibitors in American ginseng seed.Journal of Jilin Agriculture University,1994,16(2):9-14.
[16]Zhang Y J.Studies on seed dormancy mechanism of Taxus chinensis var.Mairei[D].Nanjing:Nanjing Forestry University,2007.
[17]Shang X L,Xu X Z,Fang SZ.Seed dormancy mechanism of Cyclocarya paliurus.Scientia Silvae Sinicae,2011,47(3):68-75.
[18]Nyandia C O,McPherson G R.Germination of two warm temperate oaks,Quercus emoryi and Quercus arizonica.Canadian Journal of Forest Research,1992,22(9):1395-1401.
[19]Kennedy P G,Hausmann N J,Wenk E H,Dawson T E.The importance of seed reserves for seedling performance:an integrated approach using morphological,physiological,and stable isotope techniques.Oecologia,2004,141(4):547-554.
[20]Bai K J,Guo SJ,Shi Q L.Relation between seed dormancy and embryo form,testa and inclusion of Castanea mollissima cv.‘Yanshanhong’.Journal of Southwest Forestry College,2005,25(4):106-109.
[21]Xu K,Kong Q X,Xiao SY.Studies on dormancy and germination of China chestnut seeds.Chinese Agriculture Science Bulletin,1998,14(1):24-25.
[22]Shimada T,Saitoh T.Re-evaluation of the relationship between rodent populations and acorn masting:a review from the aspect of nutrients and defensive chemicals in acorns.Population Ecology,2006,48(4):341-352.
[23]Lei N F,Su Z X,Chen JS,Guo JH.Germination inhibitors in fruit of rare and endangered Davidia involucrate.Chinese Journal of Applied and Environmental Biology,2003,9(6):607-610.
[24]Shi F H,Shen Y B,Shi JS.Study on germination inhibitor of Nanjing linden seeds.Journal of Fujian College of Forest,2007,27(3):222-225.
[25]Liu Y S.Studies on dormancy mechanism of Macrocarpium officinale seeds[D].Nanjing:Nanjing Forestry University,2008.
[26]Xu Y X.Study on Dormaney mechanism for Osmanthus americanus[D].Nanjing:Nanjing Forestry University,2008.
[27]Guo K,Li R,Werger M J A.Effect of acorn burying depth on germination,Seedling emergence and development of Quercus aliena var.acuteserrata.Acta Botanica Sinica,2001,43(9):974-978.
参考文献:
[1]李文英,王冰,黎祜琛.栎类树种的生态效益和经济价值及其资源保护对策.林业科技通讯,2001,(8):13-15.
[2]张红梅,王新杰,闫金华,白佳鸿,陆亚刚.北京市栎类植物空间分布及生长过程分析.北京林业大学学报,2010,32(增刊1):71-79.
[3]高甲荣.秦岭林区锐齿栎林水文效应的研究.北京林业大学学报,1998,20(6):31-35.
[15]黄耀阁,崔树玉,鲁歧,牛洪斌,赵春雨,韩宝瑞,杨继祥,李向高.西洋参种子抑制物质的初步研究.吉林农业大学学报,1994,16(2):9-14.
[16]张艳杰.南方红豆杉种子休眠机理的研究[D].南京:南京林业大学,2007.
[17]尚旭岚,徐锡增,方升佐.青钱柳种子休眠机制.林业科学,2011,47(3):68-74.
[20]白柯君,郭素娟,石青莲.燕山红栗种子休眠与种胚形态、种皮及内含物的关系.西南林学院学报,2005,25(4):106-109.
[21]徐凯,孔启祥,肖圣元.板栗种子休眠与萌发的研究.中国农学通报,1998,14(1):24-28.
[23]雷泞菲,苏智先,陈劲松,郭家洪.珍稀濒危植物珙桐果实中的萌发抑制物质.应用与环境生物学报,2003,9(6):607-610.
[24]史锋厚,沈永宝,施季森.南京椴种子发芽抑制物研究.福建林学院学报,2007,27(3):222-225.
[25]刘雅帅.山茱萸种子休眠机理研究[D].南京:南京林业大学,2008.
[26]许岳香.美国桂花种子休眠机理的研究[D].南京:南京林业大学,2008.
