南海西北部陆架区鱼类的种类组成与群落格局
2013-12-21王雪辉林昭进杜飞雁邱永松孙典荣王跃中黄硕琳
王雪辉 ,林昭进,杜飞雁,4,邱永松,孙典荣,王跃中,梁 新,黄硕琳
(1.中国水产科学研究院南海水产研究所,广州 510300;2.农业部南海渔业资源开发利用重点实验室,广州 510300;3.上海海洋大学海洋科学学院,上海 201306;4.厦门大学海洋与地球学院博士后流动站,厦门 361005)
南海西北部陆架区位于南海北部陆架区的边缘,地形复杂,等深线密集,同时受局地风场、黑潮入侵等因子影响,流态复杂、涡旋活跃,沿岸流与南海暖流在此交汇,夏季(6—9月)在海南岛东部沿岸及雷州半岛以东广州湾东南部一带形成琼东上升流区[1]。琼东上升流区是南海北部陆架区主要的两大上升流区之一,独特的地理位置和复杂的海流,使得该区域海洋生物资源非常丰富,形成了的著名清澜渔场和海南岛东部拖网渔场。作为南海北部陆架区的一部分,自20世纪80年代起该海域的海洋生物的研究工作就有开展,研究对象涉及鱼类和浮游植物等[2-5]。而西北部陆架区由于风生上升流的存在,使地处热带海域的琼东沿岸表层海水温度低于20℃,是我国夏季最低的表层水温,形成一年中出现两个冬天的独特生态环境[6]。然而,这一特殊区域海洋生物的专题较少开展,虽然已有研究表明夏季琼东沿岸上升流和冷涡现象对住筒虫的种类向沿岸推移及其丰度增加有一定的助长作用,但冷涡现象和上升流对海洋生物的影响及海洋生物空间分布格局状况目前尚不甚清楚[7]。
南海北部大陆架由于区域海洋生态环境的差异,近岸、浅海和深海的生物群落特点也各不相同,海洋生物区域性空间分布格局的变化,会影响海洋生物的种群动态[8]。作为海洋生态系统动力学研究的主要内容,研究鱼类群落结构及其动态变化,对了解海域生态系统的动力学机制和渔业资源的动态有着重要的意义。本研究主要以2006—2007年在南海西北部陆架区(海南岛以东海域,下同)开展的渔业资源资料,运用多元分析研究该海域鱼类的种类组成、群落结构、主要特征鱼类和群落空间格局的季节变化,以期为南海西北部陆架区海域生物群落的动态变化及演替提供基础资料,为该海域渔业资源的保护和合理利用提供科学依据。
1 材料和方法
1.1 取样
本研究所用的数据来自2006年8月、1月和2007年4月、11月在南海西北部陆架区进行的4个航次的底拖网渔业资源调查资料,分别代表夏季、冬季、春季和秋季,每航次布设调查站位16个,站位设置情况见图1。样品采样及分析均按《海洋调查规范》(GB/T 12763.6—2007)进行[9]。执行海上调查任务的调查船为北渔60011单船底拖网渔船,采样网具为404型底拖网,调查船和网具参数见表1。

表1 调查船和网具参数Table 1 The specifications of the research vessel and the fishing gear
1.2 数据处理与分析
数据分析采用非参数多变量群落结构分析方法。为减少机会种对群落结构的干扰,数据分析时去除生物量占总生物量比例<0.1%且出现频率<6.25%的种类,并对生物量进行四次方根转换,以平衡优势种和稀有种在群落中的作用。
运用转化后的数据计算站位间Bray-Curtis相似性系数,构建相似性矩阵,根据相似性矩阵,进行组平均法聚类分析和非度量多维标度(NMDS)的排序方法分析各调查站位的鱼类组成异质性[10]。由于这两种方法的互补性,在一起使用可更有效地阐明群落格局,并互相验证分析结果[11-12]。检验NMDS分析结果的优劣用胁强系数来衡量,通常认为当胁强系数<0.2时,可用NMDS的二维点图表示,其图形有一定的解释意义;当胁强系数<0.1时,可以认为是一个好的排序;当胁强系数<0.05 时,具有很好的代表性[12-14]。
应用相似性分析(ANOSIM)检验各季节不同站位间群落组成差异的显著性,群落间的相关性检验用RELATE分析进行。通过相似百分数(SIMPER)方法计算各种类对站位组内相似性和组间相异性的平均贡献率[15],并以贡献率>5%为划分特征种的标准。用BIOENV程序分析鱼类群落与水深之间的相关性[15]。
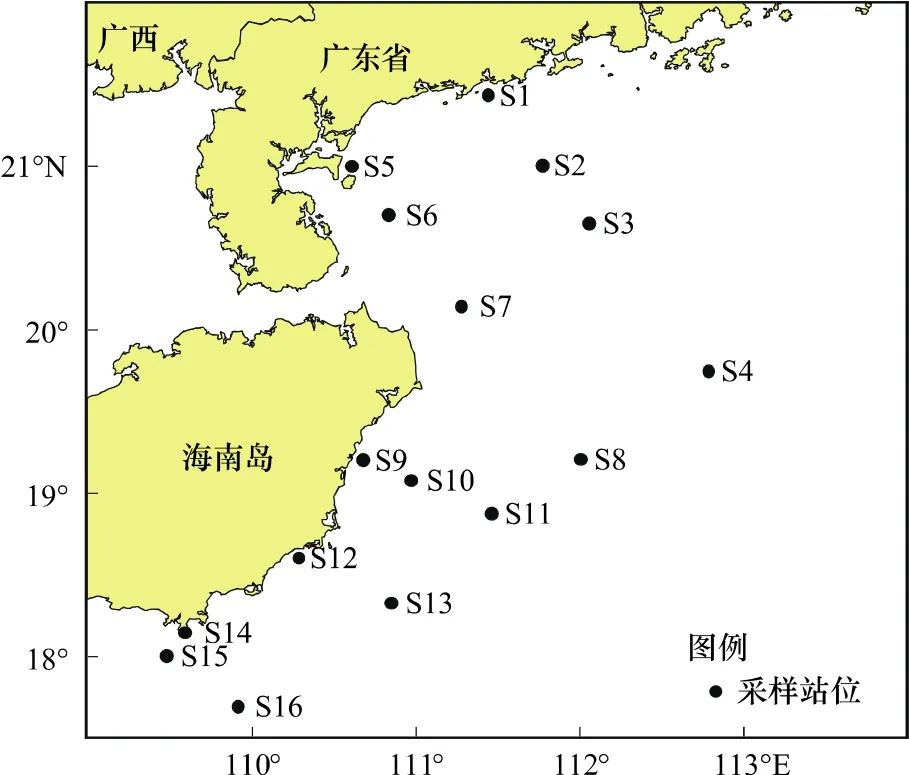
图1 采样站位示意图Fig.1 Sketch map of sampling stations in the continental shelf of northwestern South China Sea
以上分析过程使用PRIMER v5.2软件进行。图件由Surfer 8.0和PRIMER v5.2软件绘制。
2 结果
2.1 种类组成
2006—2007年4季调查共采获鱼类262种,隶属于20目102科,冬季出现种类数最多,为166种;夏季和秋季次之,分别为161种和155种;春季最少,为135种(表2)。各季均以鲈形目出现种类数最多,72种周年出现种中40种隶属于鲈形目,可见鲈形目是南海西北部陆架区海域鱼类的主要类群。四季共有种生物量占鱼类总生物量的80.34%;仅在一个季节出现的鱼类91种,其生物量占总生物量的1.62%。

表2 南海西北部陆架区鱼类的目、科和种的组成Table 2 Compositions of fish at species,family and order levels in the continental shelf of northwestern South China Sea
2.2 群落划分
图2是各采样站位按鱼类生物量进行聚类分析和NMDS排序的结果。4季NMDS分析结果的胁强系数在0.07—0.10之间,表明该图形很好地反映了群落间的相似性程度[13-14]。各季节站次聚类分析和NMDS分析结果基本一致,春季研究区域的鱼类可划分为3个群落,分别为群落Ⅰ、群落Ⅱ和群落Ⅲ;夏季和秋季可划分为4个群落,分别为群落Ⅰ、群落Ⅱ、群落Ⅲ和群落Ⅳ;冬季仅为群落Ⅰ和群落Ⅴ两个群落。群落间的ANOSIM检验表明,各季节群落间的差异极为显著(R=0.847—0.939,P<0.001,表3)。虽然各群落的站位组成和规模有较明显的季节变化,但各季重量渔获率的相似性矩阵进行Spearman相关分析结果显示,春夏、夏秋和秋冬群落间的相关系数分别为0.768、0.630和0.777,表明南海西北部陆架区鱼类群落结构较为稳定、季节变化较小(表4)。

图2 南海西北部陆架区鱼类采样站点的聚类分析和NMDS排序图Fig.2 Group average clustering and NMDS ordination of sampling stations in continental shelf of northwestern South China Sea
2.3 群落的空间分布及其优势种类
各群落的分布情况如图3所示。群落Ⅰ由3—4个测站组成,各季均有出现,稳定分布于雷州半岛东侧的粤西近岸区域;群落Ⅱ和群落Ⅲ除冬季外,其余3季均有出现。群落Ⅱ由6—9个测站组成,分布于海南岛东南沿近岸和群落Ⅰ以南的海域内,其规模和分布区域有一定的季节变化;群落Ⅲ由4个测站组成,分布于海南岛东侧、群落Ⅱ以东海域内,其规模和分布区域最为稳定;群落Ⅳ由1—3个测站组成,仅在夏、秋两季出现,分布于群落Ⅰ与群落Ⅱ之间和海南岛南部沿岸海域内,规模小且变化较大;群落Ⅴ仅在冬季出现,由13个测站组成,广泛分布于群落Ⅰ之外的大部海域内,是规模和分布区域最大的一个群落。

表3 4季节群落间的ANOSIM检验(P<0.001)Table 3 The ANOSIM test for communities in four seasons(P<0.001)

图3 南海西北部陆架区鱼类群落空间分布Fig.3 Spatial pattern of fish communities in the continental shelf of northwestern South China Sea
根据各季节鱼类群落的空间分布情况,将这些群落类型分别命名为粤西沿岸群落(群落Ⅰ)、海南岛东南沿岸群落(群落Ⅱ)、海南岛东部深海群落(群落Ⅲ)、过渡群落(群落Ⅳ)和混合群落(群落Ⅴ)。
2.3.1 粤西沿岸群落(群落Ⅰ)
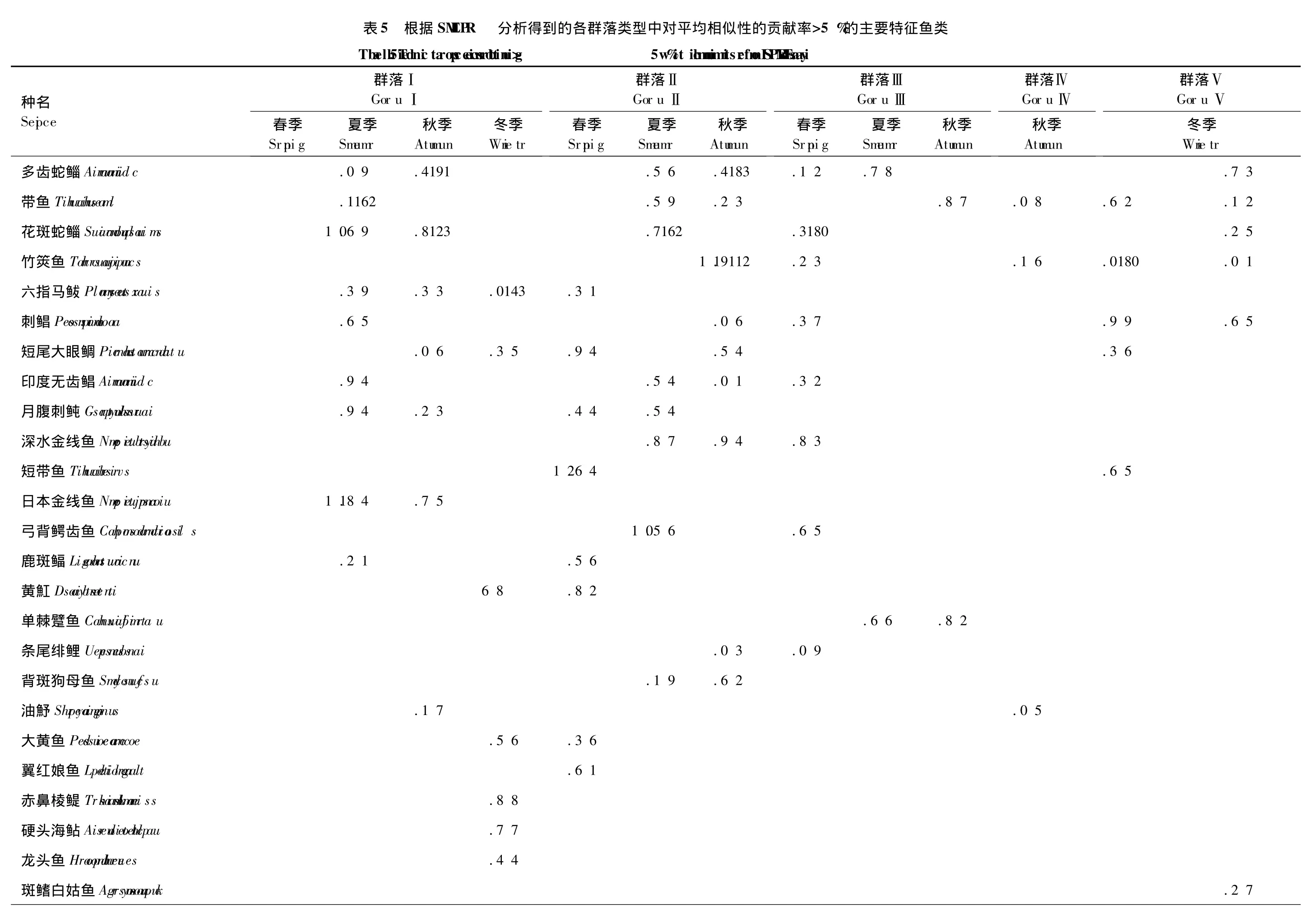
群落Ⅰ稳定分布于粤西沿岸海域,由栖息于河口、港湾和浅水的沿岸鱼类和产卵亲鱼组成。各季该群落内种类组成的相似性最高,平均相似性为60.4%,4季变化范围为55.5%—66.6%。由各季对群落的贡献率>5%为划分标准,群落Ⅰ的特征种为六指马鲅Polynemus sextarius。该群落的特征种组成较为复杂、季节变化较大,春季群落I的特征种为日本金线鱼Nemipterus japonicus、花斑蛇鲻Saurida undosquamis和刺鲳Psenopsis anomala等9种,夏季为日本金线鱼、斑鳍白姑鱼Argyrosomus pawak和油魣Sphyraena pinguis等9种,秋季为赤鼻棱鳀Thrissa kammalensis、硬头海鲇Arius leiotetocephalus和斑鳍白姑鱼等9种,冬季为六指马鲅、短带鱼Trichiurus brevis、鹿斑鲾Leiognathus ruconius和翼红娘鱼Lepidotrigla alata等10种(表5)。
粤西沿岸海域为海鲇的重要产卵场,每年冬季和春季,海鲇亲鱼集结粤西沿岸海域产卵,至5月中旬亲鱼鱼群产卵完毕后即游向深海[16]。研究表明,在冬季和春季,粤西沿岸海域聚集较多的海鲇群体,分别占冬季和春季群落Ⅰ鱼类渔获总量的33.55%和15.35%。夏季,群落Ⅰ主要以多齿蛇鲻Saurida tumbil、油魣、日本金线鱼和六指马鲅占优势,合计占该季节群落Ⅰ鱼类渔获总量的40.67%。日本金线鱼和六指马鲅不仅是夏季群落Ⅰ的主要优势种,也是群落Ⅰ四季的共有优势种。秋季,群落Ⅰ的优势种为赤鼻棱鳀和大黄鱼Larimichthys crocea,合计占该季节鱼类渔获总量的41.30%。硇洲岛附近海域曾为粤西大黄鱼的重要渔场,随着近岸捕捞力量的加大和水域环境的恶化,该海域的大黄鱼资源严重衰退[17]。近年来,南海区开始实施伏季休渔制度,严格控制捕捞业的发展,行政干预虽然使该海域的大黄鱼资源有所好转,但其产量已大不如以前。
2.3.2 海南岛东南沿岸群落(群落Ⅱ)
群落Ⅱ为海南岛东南沿岸群落,出现于春、夏和秋3个季节,以广泛分布于南海的种类为主。春、夏和秋3季群落Ⅱ种类组成的平均相似性分别为53.38%、54.58%和46.82%,平均相似性为51.8%。该群落的特征种为多齿蛇鲻、印度无齿鲳Ariomma indica和深水金线鱼Nemipterus bathybius。群落特征种组成复杂、季节变化明显,春季该群落的特征种为花斑蛇鲻、多齿蛇鲻和弓背鳄齿鱼Champsodon atridorsalis等9种,夏季特征种为竹筴鱼Trachurus japonicus、带鱼Trichiurus haumela和刺鲳等11种,秋季为多齿蛇鲻、花斑蛇鲻和竹筴鱼等8种(表5)。
2.3.3 海南岛东部深海群落(群落Ⅲ)
群落Ⅲ为海南岛东部的深海群落,出现于春、夏和秋3个季节,分布于该海域的种类主要是栖息于水深较深的或较深海的岩礁和珊瑚丛中的鱼类。春、夏和秋3季群落内种类组成的平均相似性分别为56.62%、57.52%和61.82%,平均相似性为58.8%。该群落特征种组成最为简单,没有3季对群落的贡献率>5%的种类,2季贡献率>5%的种类有带鱼和单棘躄鱼Chaunax fimbriatus。群落特征种组成的季节变化也较为明显,春季特征种为日本骨鳂Ostichthys japonicus、黄鲷Taius tumifrons和青鳞小沙丁鱼Sardinella zunasi3种,夏季为带鱼、红鳍圆鲹Decapterus russelli和无斑圆鲹Decapterus kurroides等5种(表5)。
2.3.4 过渡群落(群落Ⅳ)
群落Ⅳ为过渡群落,分布区域跨度较大,既含海南岛沿岸站位(S7和S14),也包括粤西近海站点(S2)。群落Ⅳ种类组成的平均相似性为49.11%,以黄斑鲾Leiognathus bindus、斑点马鲛Scomberomorus guttatus和二长棘鲷Parargyrops edita等7种为特征种(表5)。
2.3.5 混合群落(群落Ⅴ)
群落Ⅴ为混合群落,仅在冬季出现,群落内种类组成相似性最低,平均相似性仅为44.26%。该群落的特征种组成也最为简单,仅由斑鳍白姑鱼、单角革鲀Alutera monoceros和金线鱼Nemipterus virgatus3种组成(表5)。
类鱼征特要主%的5>率献贡的性似相均平对中型类落群各的到得析分I RPS EM据根5表3 2 5.2落ro冬ⅤuⅤt e r .7.1季in群G W 2.6Ⅳ 季um uⅣn落ro秋群G ut A 8.0 n 7季um .8秋ut uⅢ季落ro夏Ⅲr me um 8.7群G 0季pr S 2 analysi in .1 g 81 ER.3 MP春S A SI om esfr 季um 3 niti n 81 3 ncommu A秋ut .4.2 hiit %wⅡr 6 9 2 5落ro夏uⅡ季me 61群G um S .5.5.7季pr g春S in g>utinontrib季in冬W ter sc ecie sp or at n 1 3 ic季um 91 21 nd秋ut Thei A .4.8 5Ⅰle 落ro uⅠab T群G 季me r 2夏um 9 61 9 S .0.1 0.6 1季pr g春S in s c mi ua Ario m maindi l aume Sa u undosq da ri鲻蛇Tric h iurush鲻蛇名 S ci e 齿鱼斑种pe 多带花1 5.0.6 081 9.0.9 6.1 3 7.2.3 211 6 1.9.0 1 1.3341.03.39 5.3.6 s s nicu iu po xtar Trac h usja ur Po l ynemusse anomala鲅psis鱼马Psen o筴指鲳竹六刺6.3 2.3 4 1.5.0 4 4.5.5 4 4.9.4 5.36 3.0.2 4 4.9.9 u th acan cr c i Pria c usma anth Ario m maindi ophysuslunartr鲷鲳Ga s眼齿鲀大无刺尾度腹短印月3.84.97.8 Nemi p rusbathybiute鱼线金水深5.6 5.660.51 42.61 5.741.81 ssalinatridor s r u sjaponicute do so mp Tric h revi iurusb Nemi p Ch a鱼鱼线齿鱼金鳄带本背短日弓2.86.6 6 2.5.8 86 1.2 u niu at co ri ru mb fi ax un Le i netti鲾Da s ognaths tisben Ch a ya 鱼躄斑魟棘鹿黄单5.0 9.03 2.0.6 9.1 6 1.3.6 6 8.5.8 7.1 i u fusc sis Upen e nsas usbe Sy m odus uis nacrocee aalatidotrigl sakammalenis鱼n a ping osci Le p Th r鲤母y re Pseu d鱼鳀绯狗Sp h 鱼娘棱尾斑魣黄红鼻条背油大翼赤7.7 tocephaluAriu s te leio鲇海头硬7.2 4.4 eus Argy r omuspawakos Ha r donneherpo 鱼姑鱼白头鳍龙斑

?
2.4 群落与水深的关系
将鱼类群落与水深进行相关分析,春季两者之间的相关系数r=0.898、夏季r=0.852,秋季r=0.539、冬季r=0.486,相关性均达到显著水平。表明,该区域钱类群落格局与水深变化密切相关,呈明显的水深梯度变化趋势。
3 讨论
研究海域位于南海北部大陆架的西部、东侧为雷州半岛和海南岛,中东部经琼州海峡与北部湾相接,南侧与南海深海相连,地理位置特殊,受海流影响显著,环境变化较大。鱼类的分布、聚群,与其栖息环境也密切相关。现从海流变化和水深等几个方面,对研究海域鱼类群落格局的变化情况进行探讨。
3.1 群落格局变化与海流的关系
研究海域主要受到沿岸流、琼东上升流、雷州半岛东部的气旋型环流(冷涡)和南海暖流的影响,而其特殊的地理位置,使之成为沿岸流和南海暖流的交汇区[18]。南海地处东亚季风区,冬季盛行东北季风、夏季盛行西南季风,稳定强大的季风对南海环流的影响非常显著[19]。因此,南海海流的季节变化也较为复杂。鱼类的分布受栖息环境的影响,研究区域的鱼类,春季分为群落Ⅰ、群落Ⅱ和群落Ⅲ3个群落;夏和秋季分为群落Ⅰ、群落Ⅱ、群落Ⅲ和群落Ⅳ4个群落;冬季分为群落Ⅰ和群落Ⅴ两个群落,研究海域复杂的海流运动,是鱼类群落格局发生较大变化的主要原因之一。
南海北部沿岸海域终年受沿岸流的影响,沿岸流受到雷州半岛的阻挡后,在粤西外侧风生海流的影响下,在雷州半岛东部水域(20°20'—21°10'、110°50'—112°)形成一个逆时针的环流(环流),引发底层水涌升[18,7]。冷涡终年存在,夏强冬弱。独特而相对稳定的环境,使该区域周年栖息着大量的咸淡水鱼类,从而形成最为稳定的粤西沿岸群落——群落Ⅰ。海南岛的东南外海的南海暖流,沿大陆架流向东北,其流向稳定、终年不变[18,20]。在南海暖流控制的区域主要栖息着深水鱼类,从而形成了海南岛东部深海群落——群落Ⅲ。冬季,这支东北向的逆风暖流势力强劲,在海南岛东南方相当稳定[21]。在其影响下研究海域的鱼类,除稳定分布在雷州半岛东北侧的群落I外,沿岸型和深水性种类均集结为混合性群落——群落Ⅴ。春、夏和秋季,沿岸流势力增强,南海暖流势力则有所减弱。在季风和海底地形的影响下,南海暖流流经的海南岛东侧形成强劲的近岸上升流,在此区域鱼类形成海南岛东南沿岸群落——群落Ⅱ。此外,在沿岸流势力强劲的夏、秋两季,沿岸流和上升流的交汇处,粤西和北部湾海流相通的小范围海域内流向发生变化[20]。独特的海流和栖息环境,鱼类在此形成了范围较小的过渡群落——群落Ⅳ。鱼类的聚群和空间分布与栖息环境密切相关,研究海域受海流影响显著,海流的季节性变化也非常明显,直接影响着栖息于该区域的鱼类群落的聚合,而该海域鱼类群落格局的变化也很好地反映了此处海流的变化与消长状况。
3.2 群落格局与水深的关系
研究区域位于南海北部大陆架边缘,等深线密集、水深变化梯度明显。Odum和邱永松研究均表明,水深、温度和盐度等环境因子是游泳动物群集的主要限制因素,而水深是影响鱼类群落结构变化的最主要因素[22,3,23]。水深引起群落变化的不是水深本身,而是与水深变化有伴随关系的底层水温和盐度的变化[3]。不同深度的海域具有不同的环境,环境因素对鱼类分布起限制作用,使不同的水系和水深常成为不同种类的主要分布区,从而形成了鱼类群落。本文对鱼类群落与水深的相关分析也反映出群落与水深的变化密切相关,从群落的分布格局来看,研究区域的群落有明显的沿水深梯度变化趋势,呈较明显的沿岸、近岸和远岸的分布规律。
[1]Jing Z Y,Qi Y Q,Hua Z L.Numerical study on summer upwelling over northern continental shelf of South China Sea.Journal of Tropical Oceanography,2008,27(3):1-8.
[2]Fei H N,He B Q,Chen G M.The regional and seasonal variations of diversity and dominant species of demersal fish communities in continental shelf of northern Nanhai.Journal of Fisheries of China,1981,5(1):1-20.
[3]Qiu Y S.The regional changes of fish community on the northern continental shelf of South China Sea.Journal of Fisheries of China,1988,12(4):303-311.
[4]Jia X P,Li C H,Qiu Y S.Assessment and Sustainable Utilization of Marine Fisher Resources in Guangdong Province.Beijing:Ocean Press,2005:1-322.
[5]Cai W G,Li C H,Jia X P,Lin Q,Gan JL.Multivariate analysis and evaluation on the community structure of phytoplankton in west Guangdong waters.Acta Hydrobiologica Sinica,2007,31(2):155-161.
[6]Han W Y,Wang M B,Ma K M.On the lowest surface water temperature area of China Sea in summer-the upwelling along the east coast of Hainan Island.Oceanologia et Limnologia Sinica,1990,21(3):267-275.
[7]Li K Z,Yin J Q,Huang L M,Chen Q C,Lin Q.Taxonomic identification and abundance distribution of fritillarians in the northwest continental shelf of South China Sea.Acta Oceanologica Sinica,2010,32(5):76-86.
[8]Shen G Y,Huang L F,Guo F,Shi B Z.Marine Ecology.3rd ed.Beijing:Science Press,2010:1-85.
[9]Standardization Administration of China.GB/T 12763.6—2007 Specifications for oceanographic(survey-Part 6:Marine biological survey).Beijing:Standards Press of China,2007.
[10]Field JG,Clarke K R,Warwick RM.A practical strategy for analysing multispecies distribution patterns.Marine Ecology Progress Series,1982,8:37-52.
[11]Brazner JC,Beals EW.Patterns in fish assemblages from coastal wetland and beach habitats in Green Bay,Lake Michigan:a multivariate analysis of abiotic and biotic forcing factors.Canadian Journal of Fisheries and Aquatic Sciences,1997,54(8):1743-1761.
[12]Li SF,Chen JH,Yan L P.The spatial pattern of the fish assemblage structure in the mid-southern East China Sea.Acta Oceanologica Sinica,2005,27(3):110-118.
[13]Khalaf M A,Kochzius M.Changes in tropic community structure of shore fishes at an industrial site in the Gulf of Aqaba,Red Sea.Marine Ecology Progress Series,2002,239:287-299.
[14]Ma Z Y,Liu H,Wang H Q,Wang SQ.Multivariate analysis of community structure on macrobenthos.China Environmental Science,1997,17(4):297-300.
[15]Clarke K R,Warwick R M.Change in Marine Communities:An Approach to Statistical Analysis and Interpretation.2nd ed.Plymouth:PRIMPER-E,2001.
[16]Chen Z C,Liu JX.Commercial fishes of South China Sea.Guangzhou:Guangdong Science and Technology Press,1982:60-66.
[17]Yang R H.Status of Pseudosciaena crocea resource and suggestions of its protection around Naozhou Island.Fisheries Science and Technology,1995,(3):1-4.
[18]Yu M G,Liu J F.Current system and circulation pattern in the South China Sea.Marine Forecasts,1993,10(2):13-17.
[19]Yang H J,Liu Q Y.A summary on ocean circulation study of the South China Sea.Advance in Earth Sciences,1998,13(4):364-368.
[20]Yang SY,Bao X W,Chen CS,Chen F.Analysis on characteristics and mechanism of current system in west coast of Guangdong Province in the summer.Acta Oceanologica Sinica,2003,25(6):1-8.
[21]Guan B X.Some features of the temporal and spatial distributions of the“counter-wind”current in northern South China Sea in winter.Oceanologia Et Limnologia Sinica,1985,16(6):429-438.
[22]Odum E.Fundamentals of Ecology//Sun R Y,Qian G Z,Lin H R,Zhu J,Li Z Y,Lu H J,translateds.3rd ed.Beijing:People Education Press,1981:320-346.
[23]Colvocoresses JA,Musick JA.Species associations and community composition of middle Atlantic Bight continental shelf demersal fishes.Fishery Bulletin,1984,82(2):295-313.
参考文献:
[1]经志友,齐义泉,华祖林.南海北部陆架区夏季上升流数值研究.热带海洋学报,2008,27(3):1-8.
[2]费鸿年,何宝全,陈国铭.南海北部大陆架底栖鱼群聚的多样度以及优势种区域和季节变化.水产学报,1981,5(1):1-19.
[3]邱永松.南海北部大陆架鱼类群落的区域性变化.水产学报,1988,12(4):303-311.
[4]贾晓平,李纯厚,邱永松.广东海洋渔业资源调查评估与可持续利用对策.北京:海洋出版社,2005:1-322.
[5]蔡文贵,李纯厚,贾晓平,林钦,甘居利.粤西海域浮游植物群落结构特征的多元分析与评价.水生生物学报,2007,31(2):155-161.
[6]韩舞鹰,王明彪,马克美.我国夏季最低表层水温海区-琼东沿岸上升流区的研究.海洋与湖沼,1990,21(3):267-275.
[7]李开枝,尹健强,黄良民,陈清潮,林强.南海西北部陆架区住筒虫属(Fritillaria)的种类描述及其丰度分布.海洋学报,2010,32(5):76-86.
[8]沈国英,黄淩风,郭丰,施并章.海洋生态学(第三版).北京:科学出版社,2010:1-85.
[9]中国国家标准化管理委员会.GB/T 12763.6—2007海洋调查规范(第6部分海洋生物调查).北京:中国标准出版社,2007.
[12]李圣法,程家骅,严利平.东海中南部鱼类群聚结构的空间特征.海洋学报,2005,27(3):110-118.
[14]马藏允,刘海,王惠卿,王世权.底栖生物群落结构变化多元变量统计分析.中国环境科学,1997,17(4):297-300.
[16]陈再超,刘继兴.南海经济鱼类.广州:广东科学技术出版社,1982:60-66.
[17]杨日晖.硇洲岛附近大黄鱼资源状况及资源保护的建议.水产科技,1995,(3):1-4.
[18]俞慕耕,刘金芳.南海海流系统与环流形势.海洋预报,1993,10(2):13-17.
[19]杨海军,刘秦玉.南海海洋环流研究综述.地球科学进展,1998,13(4):364-368.
[20]杨士瑛,鲍献文,陈长胜,陈菲.夏季粤西沿岸流特征及其产生机制.海洋学报,2003,25(6):1-8.
[21]管秉贤.南海北部冬季逆风海流的一些时空分布特征.海洋与湖沼,1985,16(6):429-438.
[22]奥德姆.生态学基础//孙儒泳,钱国桢,林浩然,译.3版.北京:人民教育出版社,1981:320-346.
