海岸带地理特征对沉水植被丰度的影响
2013-12-20吴明丽李叙勇陈年来
吴明丽,李叙勇 ,陈年来
(1.甘肃农业大学资源与环境学院,兰州 730070;2.中国科学院生态环境研究中心城市与区域生态重点实验室,北京 100085;3.美国Smithsonian环境研究中心,美国马里兰州 21037)
沉水植被(SAV)是指植被体全部位于水面以下的大型水生草本植物,其通气组织发达,各部分都可吸收水分和养料,皮层细胞含有叶绿素,可进行光合作用。沉水植被为水生生物提供了生存环境、对抑制生物性和非生物性悬浮物、改善水下光照和溶解氧条件、净化水质等具有重要作用[1-2]。沉水植被在水生生态系统中的作用越来越受到人们的重视。然而,随着水体污染的加剧和水体富营养化进程的加快,自20世纪60年代以来,沉水植被在世界范围内出现了大面积的减少。光照衰减及营养物和悬浮物的增加使沉水植被的生长环境极度恶化,有很多学者和专家在水质和光照衰减对沉水植被的影响方面做了大量的工作,附生植被、总悬浮颗粒物、叶绿素和营养物等往往是考虑最多的因素[3-5]。近年来,景观分析开始应用于海岸带地理特征对水生生物直接或间接的影响上[6-9]。流域发展可以改变河口的物种多样性、改变食物网以及底栖生物群落组成结构,流域土地利用类型是健康河口是否退化的一个重要指示器[10-11]。通常,沉水植被生长在浅海岸地带的溪流和港湾中,这些浅滩是陆地和海洋的过渡地带,流域通过影响河口水质影响着沉水植被丰度,而河口带的沉积物、来自海洋方向的浪潮等则直接或间接影响着沉水植被的丰度。本文是在前期研究的基础上[7],增加了水体高分辨率海浪高度、沉积物和水体深度等特征数据,并增加了近几年(2004—2009年)航片解译的沉水植被丰度数据,系统研究了海岸带地理特征(流域土地利用类型、河口宽度、海岸线分形维度、流域面积与河口面积比、高分辨率平均波浪高度和沉积物等)对沉水植被的长期分布(1984—2009年)产生的可能影响,从而为制定区域尺度上沉水植被的保护和恢复提供科学依据。
1 研究区概况与数据源
1.1 研究区概况
切斯比克海湾位于美国东部的马里兰州和弗吉尼亚州海岸带(图1)[12]。海湾水面面积5720 km2,流域面积16.6万km2,宽度最窄处5.5 km,最宽处56 km,平均深度6.4 m;整个流域包括150条大的支流,其中最长的最主要的河流是萨斯奎汉纳河,南北纵贯宾夕法尼亚州,切斯比克海湾50%以上的淡水来自此河。切斯比克海湾海岸线蜿蜒曲折,长达6440 km,是美国重要的渔业生产基地之一,既有来自河流的充沛淡水,又有来自大西洋的咸水,水深较浅,光照条件好,适合各类水生生物生长。在海湾内生长的动植被极为丰富,鱼类295种。贝类45种,沉水植被等各种植被达2700多种。近几十年来,切斯比克海湾资源消耗及水体污染严重,生态环境条件恶化,沉水植被出现了大面积的减少。
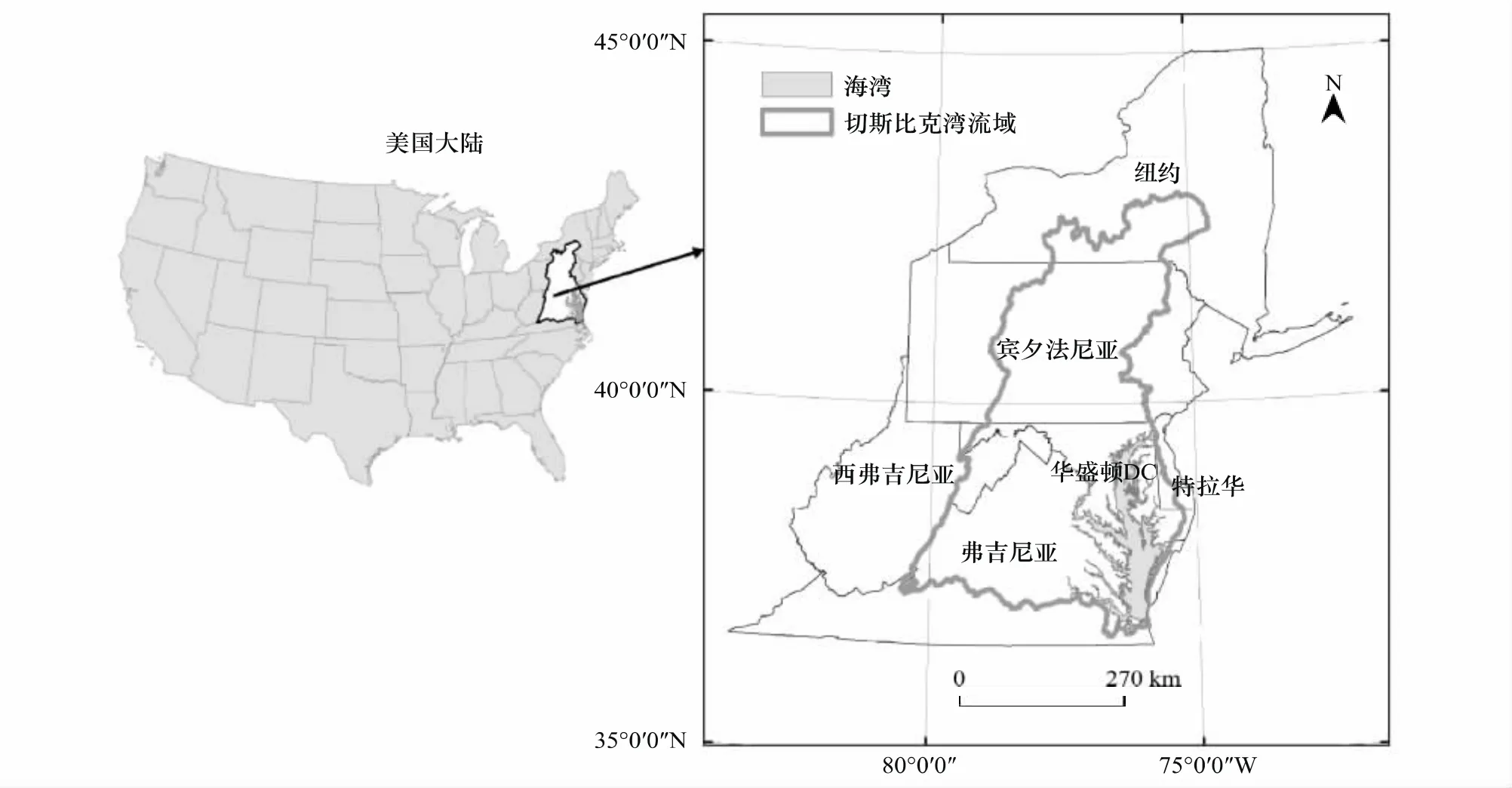
图1 切斯比克海湾流域Fig.1 The location of Chesapeake Bay Watershed
1.2 数据来源与整理
文章选取切斯比克海湾128个河口及其所在流域作为分析目标(图1)。沉水植被丰度数据来自美国弗吉尼亚海洋研究所1984—2009年的高分辨率(1∶24000)航片解译资料[13],运用ArcGIS软件提取得到,由于某些河口多数没有沉水植被生长,以及两个河口为人工栽培的沉水植被,剔除这些河口之后,实际选取了99个河口来做分析。沉水植被丰度计算公式为:
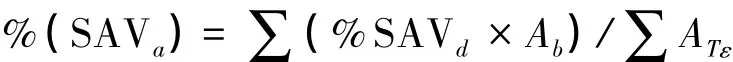
式中,%(SAVa)为某个河口的沉水植被丰度;%SAVd为在一个沉水植被生长的基床监测到的沉水植被密度;Ab为沉水植被生长的基床数量;ATε表示估计到的沉水植被在一个基床上能够恢复生长的最大丰度。流域土地利用类型根据TM影像解译分为4种类型:建设用地、农业用地、混合用地和林地。盐度分区为:淡水区、低盐度区、中盐度区和高盐度区。用ArcGIS软件计算出了河口宽度、海岸线分形维度、流域面积与河口面积比等数据,各河口水体深度来自于美国环境署切斯比克海湾计划办公室的海底水体分布图,河口水体高分辨率(1∶24000)沉积物(由砂粒、粘粒和粉粒组成)及海浪分布图由美国马里兰州自然资源部提供。主要采用SPSS16.0和R软件来进行数据的分析和处理。
2 结果与讨论
2.1 沉水植被在不同盐度分区及不同土地利用类型下的丰度分布
流域土地利用类型是底栖生物群落是否退化的一个重要的指示器[11]。盐分浓度的差异是沉水植被丰度空间分布变异的重要因子。频繁的盐度波动是造成河口沉水植被衰退的主要原因之一[14]。由图2和图3可以看出,所有河口地区及不同盐度分区,沉水植被在不同的土地利用类型下的丰度分布差异显著,差异性随盐度区域的不同而不同;不同土地利用类型分区下,沉水植被在不同盐度区域下的丰度分布也显著,差异性随土地利用类型的不同而不同。这说明土地利用类型和盐度是影响区域沉水植被丰度变化的重要因素。流域内不同土地利用类型对向河口输送的营养物、沉积物及其他污染物的影响不同,引起的水质条件的变化也不相同,进而对沉水植被丰度变化的影响也有所差异。如图3,沉水植被在建设用地丰度分布相对较低,这是因为建设用地会带来更多的污染物质的排放,如市政生活污水、草坪施肥和工业活动等排放的污染物输入到河口中,引起水体环境的改变,水体富营养化以及水体浑浊度增加,减少光照的利用,进而影响沉水植被丰度的分布;也因为建设用地有较大比例的不透水地面,使悬浮物和营养物更容易直接流入水体,使沉水植被在这种土地利用类型下丰度较低。在淡水区和低盐度区(图2),沉水植被在林业用地上的分布较低(图3),这可能是因为在淡水区和低盐度区生长的沉水植被有色可溶性有机质(CDOM)引起的光穿透性降低更为敏感,而在切斯比克海湾,降雨对流域森林土壤腐殖质表层的冲刷携带大量的CDOM进入近海浅水区域[3],是引起光穿透性降低,从而影响沉水植被生长的重要原因。沉水植被在中盐度区域各个土地利用类型上的分布较低(图2),这说明盐度胁迫对水生植被的生长具有很大的影响,也有研究表明,盐度是影响沉水植被种子萌发的环境因子之一[15];盐度能够改变抑制川蔓藻和蓖齿眼子菜的平均比生长速率[14],是影响沉水植被丰度分布的重要因素。
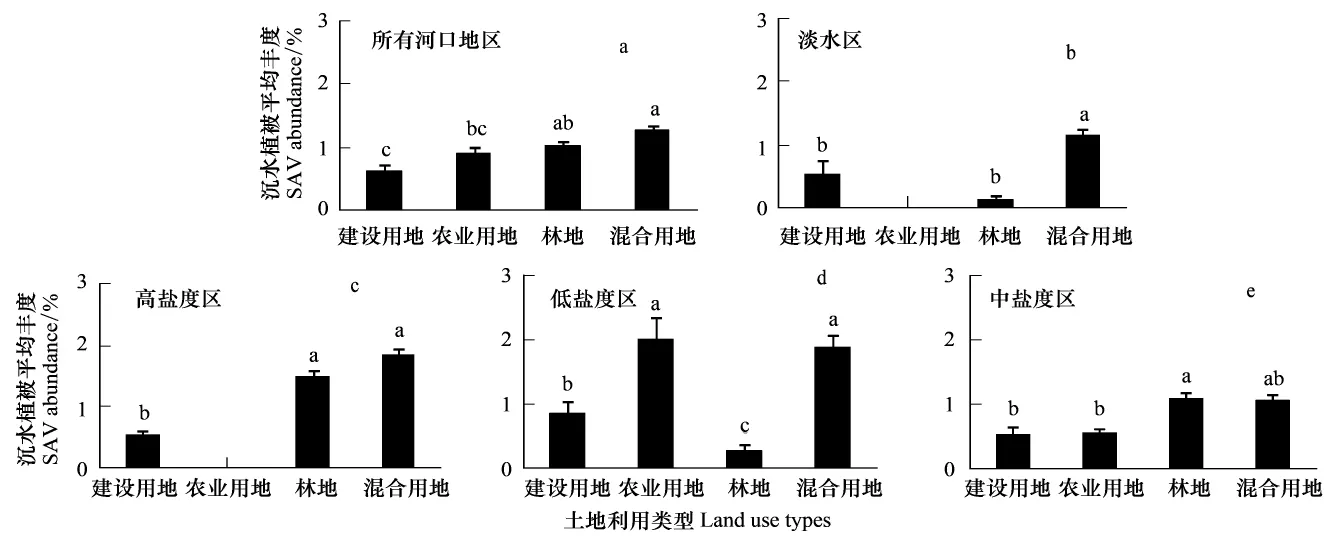
图2 所有河口及不同盐度分区,沉水植被在不同土地利用类型下的丰度分布Fig.2 SAV abundance in sub-estuaries with different dominant watershed land use types for each salinity regime
2.2 海岸带地理特征与沉水植被丰度的相关性分析
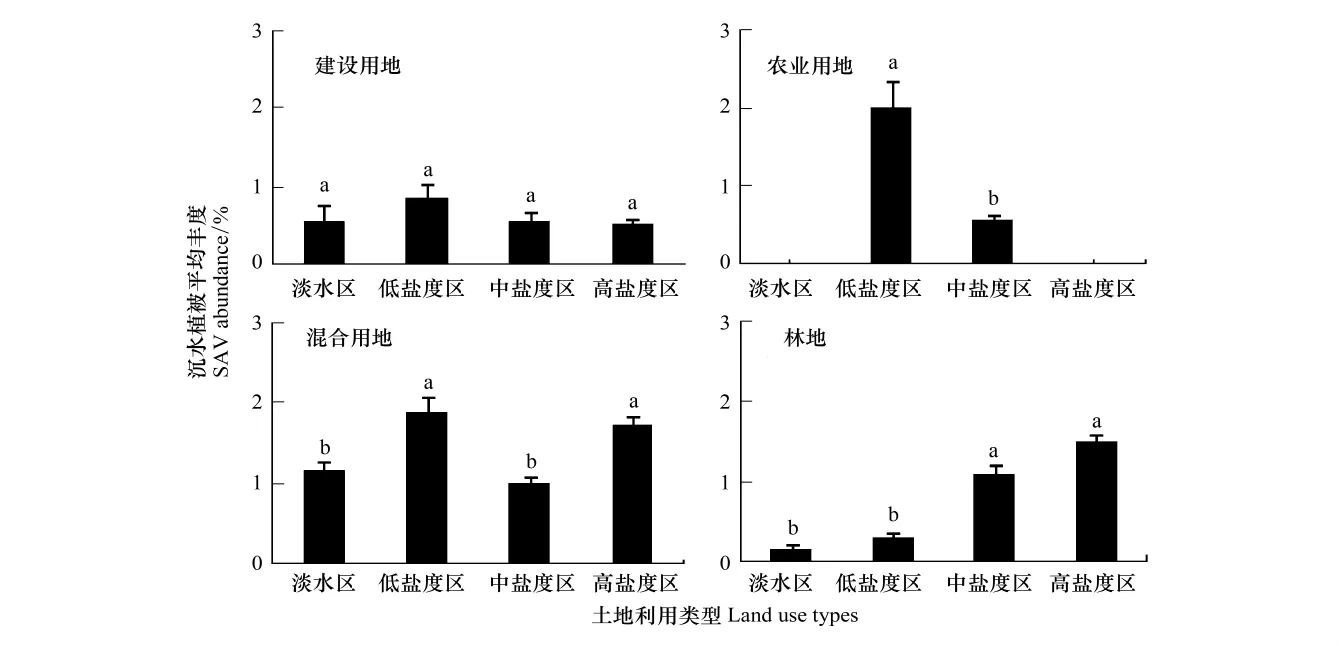
图3 不同土地利用类型分区下,沉水植被在不同盐度地区的丰度分布Fig.3 SAV abundance in sub-estuaries with different salinity regimes for each dominant watershed land use type
由图4可以看出,(1)沉水植被丰度与建设用地(%)的关系(图4):沉水植被多数生长在建设用地为0—20%的范围内,呈微弱负相关关系;但在>20%的建设用地上,沉水植被丰度随着建设用地的增加出现了大幅下降的趋势。这是因为建设用地上多为排污系统、工业活动等会带来更多的污染物,直接或者间接的排放到河流中,造成河水水质恶化,从而影响到沉水植被生长,用于发展建设的用地越高对沉水植被的影响也就越大,丰度就会越来越少。(2)沉水植被丰度与河口宽度(km)关系(图4):沉水植被多数生长在河口宽度为0—3(km)的范围内,两者呈正相关关系。相比较其他特征因子,沉水植被丰度与河口宽度的相关性相对较高,随着河口宽度的增加,沉水植被丰度值也相对增加。这是因为河口宽度的大小对污染物、悬浮物的稀释具有很大影响,较宽的河口,流动性较好,与外界交换水体的能力就越大,稀释污染物的能力就越高,河水水质就会相对较好,适于沉水植被生长的环境条件也就较好。(3)沉水植被丰度和流域与河口面积比关系(图4):沉水植被多数生长在流域与河口面积比为0—20的范围内,沉水植被丰度与其呈负相关关系;但是在二者面积比大于20时,沉水植被随着面积比的增加出现大幅下降,呈较强的负相关关系,这表明,单位面积海湾所连接的流域陆地面积越大,适合沉水植被生长的条件就越低,沉水植被丰度就越低。选取的切斯比克海湾99个河口均为比较小的半封闭浅海浅滩地区,在这些区域营养物及悬浮物相比较于大的河口及开放水域来说受河道的冲刷和稀释就会低很多,因此把流域和河口面积比这一关系作为一个考虑因素。(4)沉水植被丰度与海岸线分形维度的关系(图4):沉水植被丰度与海岸线分形维度呈微弱极显著正相关关系。海岸线分形维度越高,沉水植被丰度就越高。这是因为,海岸线越是曲折复杂,就越能形成一些易于沉水植被生长的狭小港湾,更多的营养物沉淀下来,且避免受到大风大浪的冲刷,沉水植被丰度相对较高。(5)沉水植被丰度与高分辨率平均波浪高度的关系(图4):沉水植被丰度与平均波浪也是呈微弱极显著正相关关系。波浪对沉水植被具有正反两方面的影响:一方面平均波浪越高越能带来一部分营养物,供植被生长,波浪扰动也能使粘附在沉水植被叶片表面的悬浮物减少,提高水体透明度,易于光照达到沉水植被叶片表面,进行光合作用;另一方面波浪扰动也能使一部分悬浮物黏附在叶片表面,对光照产生遮挡,影响其光合作用[7,16]。(6)沉水植被丰度与沉积物的关系(图4):沉水植被丰度与沉积物呈微弱正相关关系。沉积物可以带来一定的营养物质,促进沉水植被生长;但沉积物粘附在叶片上,也能使叶片光合作用减少,不利于沉水植被生长。(7)沉水植被丰度与其平均生长水体深度的关系(图4):沉水植被丰度与其生长平均深度呈微弱负相关关系。如图3沉水植被多数生长在-4—0(m)浅海浅滩地区,在这一范围内,沉水植被丰度值几乎没有变化,说明沉水植被在此深度内生长良好。
2.3 综合考虑多种相关特征因子与沉水植被丰度变化的关系
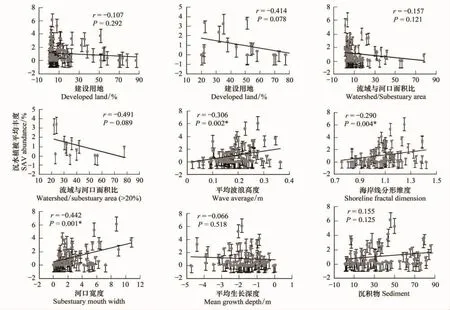
图4 海岸带地理特征与沉水植被丰度相关关系图Fig.4 Correlations of SAV abundance with a single variable of coastal geographical characteristics
图5为各个特征因子对沉水植被丰度影响的分类回归树图,这个几个因子共解释了沉水植被丰度空间变异的60%以上。可以看出,不同特征因子对沉水植被丰度影响的贡献值不同,影响大小不同。其中流域和河口面积比排在树的最顶端,当流域与河口面积比小于3.509时,有13个河口,沉水植被平均丰度为2.458%;海岸线分形维度排在树的第2位,当分形维度小于0.985时,有 26个河口,沉水植被平均丰度为0.3196%;排在树的第3位为盐度因素,淡水区和中盐度为一分支,低盐度和高盐度为另一分支;排在树的最后分支为流域与面积比和平均波浪高度,其中当流域与河口面积比≥5.9时,有21个河口,沉水植被平均丰度为0.3448%,当面积比<5.9时,有14个河口,沉水植被平均丰度为1.197%;当平均波浪高度<0.1869时,有13个河口,沉水植被平均丰度为1.15%,当波浪高度≥0.1869时,有 12个河口,沉水植被平均丰度为2.372%。以上是对切斯比克海湾99个河口做的一个综合分析。其中,流域与河口面积比对沉水植被丰度的影响是最大的,与图3相对应,流域和河口面积比值越大,适合沉水植被生长的条件就越低,沉水植被丰度相应减少;海岸线分形维度对沉水植被丰度的影响也较大,分形维度越大,越是能形成一些有利于沉水植被生长的港湾[17-18],且避免受到风浪冲刷侵蚀,沉水植被丰度也就越高。

图5 海岸带地理特征对沉水植被丰度影响的回归树分析Fig.5 The classification and regression tree model predicting SAV abundance by coastal geographical characteristics
3 结论
(1)所有河口地区及不同盐度区域,沉水植被在不同的土地利用类型下的丰度分布差异显著,差异性随盐度区域的不同而不同;沉水植被在不同盐度区域的丰度分布差异也显著,差异性随土地利用类型分区的不同而不同。
(2)单个特征因子与沉水植被丰度之间的相关关系微弱。其中沉水植被丰度与河口宽度、海岸线分形维度和高分辨率平均波浪高度相关关系相对较高,且具有极强的统计学意义(P<0.01)。
(3)不同海岸带地理特征因子对沉水植被丰度的影响不同;同一因子不同范围内对沉水植被丰度影响的大小不同,如>20%的建设用地和>20%的流域与河口面积比所做的分析中,沉水植被则出现大幅减少的状况。
(4)不同特征因子对沉水植被丰度的影响不同。分析的99个河口和25a(1984—2009年)时间内,流域和河口面积比是影响沉水植被丰度最大的因素,排在分类回归树的最顶端,依次为海岸线分形维度和盐度等因素。
[1] Wang H,Pang Y,Liu S B,Ma X.Research progress on influencing of environmental factors on the growth of submersed macrophytes.Acta Ecologica Sinica,2008,28(8):3958-3968.
[2] Van Donk E,van de Bund W.Impact of Submerged macrophytes including charophytes on phyto and zooplankton communities:allelopathy versus other mechanisms.Aquatic Botany,2002,72(3/4):267-274.
[3] Gallegos C L.Calculating optical water quality targets to restore and protect submersed aquatic vegetation:Overcoming problems in partitioning the diffuse attenuation coefficient for photosynthetically active radiation.Estuaries,2001,24(3):381-397.
[4] Kemp W M,Batleson R,Bergstrom P,Carter V,Gallegos C L,Hunley W,Karrh L,Koch E W,Landwehr J M,Moore K A,Murray L,Naylor M,Rybicki N B,Stevenson J C,Wklcox D J.Habitat requirements for submerged aquatic vegetation in Chesapeake Bay:water quality,light regime,and physical-chemical factors.Estuaries,2004,27(3):363-377.
[5] Pérez G P,Llames M E,Lagomarsino L,Zagarese H.Seasonal variability of optical properties in a highly turbid lake(Laguna Chascomús,Argentina).Photochemistry and Photobiology,2011,87(3):659-670.
[6] King R S,Hines A H,Craige F D,Grap S.Regional,watershed and local correlates of blue crab and bivalve abundances in subestuaries of Chesapeake Bay,USA.Journal of Experimental Marine Biology and Ecology,2005,319(1/2):101-116.
[7] Li X Y,Weller D E,Gallegos C L,Jordant T E,Kim H C.Effects of watershed and estuarine characteristics on the abundance of submerged aquatic vegetation in Chesapeake Bay subestuaries.Estuaries and Coasts,2007,30(5):840-854.
[8] Rodriguez W,August P V,Wang Y Q,Paul J F,Gold A,Rubinstein N.Empirical relationships between land use/cover and estuarine condition in the northeastern United States.Landscape Ecology,2007,22(3):403-417.
[9] Latimer J S,Charpentier M A.Nitrogen inputs to seventy-four southern New England estuaries:application of a watershed nitrogen loading model.Estuarine,Coastal and Shelf Science,2010,89(2):125-136.
[10] Hale S S,Paul J F,Heltshe J F.Watershed landscape indicators of estuarine benthic condition.Estuaries,2004,27(2):283-295.
[11] Leight A K,Slacum W H,Wirth E F,Fulton M H.An assessment of benthic condition in several small watersheds of the Chesapeake Bay,USA.Environmental Monitoring and Assessment,2011,176(1/4):483-500.
[12] Tian D,Li X Y,Weller D E,Bai Z K.Impacts of land use and impervious surface on stream flow metrics in Chesapeake Bay watershed.Journal of Natural Resources,2011,26(6):1012-1020.
[13] http://www.vims.edu/bio/sav.
[14] Wang W H,Ji M,Xue Y W.Response mechanism of ruppia maritima and potamogeton pectinatus to salinity in reclaimed wastewater.Journal of Tianjin University,2007,40(7):804-810.
[15] Yu L Y,Jiang L Z.Effects of different concentration of NaCl solution on seed germination of V.spiralis,V.spinulosa and Ottelia alismoides.Anhui Agricultural Science Bulletin,2008,14(7):77-79.
[16] Clarkes S M.Seagrass-sediment dynamics in Holdfast Bay:Summary.Safish,1987,11(2):4-10.
[17] Chaplin G I,Valentine J F.Macroinvertebrate production in the submerged aquatic vegetation of the Mobile-Tensaw Delta:effects of an exotic species at the base of an estuarine food web.Estuaries and Coasts,2009,32(2):319-332.
[18] Lafabrie C,Major K M,Major C S,Cebrián J.Arsenic and mercury bioaccumulation in the aquatic plant,Vallisneria neotropicalis.Chemosphere,2011,82(10):1393-1400.
参考文献:
[1] 王华,逄勇,刘申宝,马璇.沉水植物生长因子研究进展.生态学报,2008,28(8):3958-3968.
[12] 田迪,李徐勇,Weller D E,白中科.土地利用及不透水地表对河流流量的影响.自然资源学报,2011,26(6):1012-1020.
[14] 王卫红,季民,薛玉伟.川蔓藻和蓖齿眼子菜对再生水中盐度的响应机制.天津大学学报,2007,40(7):804-810.
[15] 袁龙义,江林枝.不同盐度对苦草、刺苦草和水车前种子萌发的影响研究.安徽农学通报,2008,14(7):77-79.
