人工巢箱条件下两种山雀鸟类的同域共存机制
2013-12-25殷江霞刘子成万冬梅
李 乐,张 雷,殷江霞,刘子成,刘 鹤,万冬梅
(辽宁大学生命科学院,辽宁省动物资源与疫病防治重点实验室,沈阳 110036)
生态位是生态学中的一个重要概念,主要指在自然生态系统中一个种群在时间和空间上的位置及其与相关种群之间的关系[1]。由于竞争的存在,两个生态学上相似的物种不能占有相同的生态位,而是以某种方式彼此取代,每种各具不同的食性或其他生活方式上的特点[2]。在长期的自然进化过程中,为了最大限度地提高自身的适合度,物种形成了一系列的繁殖对策和生存对策[3]。目前,对同域分布而又生态需求上十分相似物种共存机制的研究渐渐成为研究热点,涉及繁殖行为[4-6],食性[7-8]和觅食地生境选择[9-13]等方面。Dhondt通过对欧洲大山雀(Parusmajor)和青山雀(Cyanistescaeruleus)的比较研究发现,两者的食性差异不大,但其竞争强度在质量差的生境中大于质量好的生境,而且竞争对大山雀的影响程度要大于青山雀[8];崔鹏等对繁殖季节红腹角雉(Tragopantemminckii)和血雉(Ithaginiscruentus)的研究表明,红腹角雉活动区内的箭竹林面积比例显著小于血雉活动区, 针阔混交林面积比例显著大于血雉,觅食生境的差异是这两种雉类能够同域分布的基础条件之一[11];赵亮等对角百灵(Eremophilaalpestris)和小云雀(Alaudagulgula)巢址选择的研究表明,这两种百灵鸟类在巢址的选择上存在着一种代价和利益的权衡关系,巢址选择上的分化是它们能够同域共存的基础[13]。如前文所述,种间竞争会导致鸟类对自身的生存策略进行多方面调整。目前对鸟类共存机制的研究多从单一因素出发,这并不能全面地揭示鸟类共存的原因。因此,将多种因素结合起来分析将具有十分重要的意义。
大山雀和杂色山雀(Parusvarius)同属雀形目,山雀科的小型森林洞巢鸟类。大山雀属于广布种,主要分布在中国、非洲西北部摩洛哥、突尼斯、阿尔及利亚、欧洲、中亚、西伯利亚、远东、朝鲜、日本、阿富汗、巴基斯坦、印度、中南半岛、印度尼西亚。全球共计31个亚种,我国分布有6个亚种。主要栖息于低山和山麓地带的次生阔叶林和针阔混交林中,也出入人工林和针叶林中[14]。杂色山雀属狭布种,为东亚特有鸟类,主要分布在中国、日本和朝鲜半岛,全球曾有9个亚种,其中大东岛亚种P.v.orii于1930年灭绝,现在全球只剩8个亚种。我国仅分布有2个亚种,分别是指名亚种P.v.varius和台湾亚种P.v.castaneoventris,指名亚种主要分布在辽宁中、东、南部山区和毗邻辽东山区的吉林省部分山区[14-15];台湾亚种被台湾当地称做赤腹山雀,是台湾特有亚种,在台湾岛呈不连续分布,主要栖息在低于1000 m的中低海拔的阔叶林中,台中的八仙山区是杂色山雀分布比较集中的地区。
在辽宁省东部山区,大山雀与杂色山雀的分布区域基本重叠,而且同域繁殖,均是从3月中下旬开始配对活动,至7月下旬繁殖结束,2009年5月调查时发现两种鸟类的最近繁殖巢间距仅为15 m左右,这为研究不同鸟类间的共存机制提供了便利。本文于2009—2011年3—7月,以悬挂人工巢箱的方法招引杂色山雀和大山雀,通过比较这两种鸟类在繁殖时间、繁殖参数和巢址选择上的差异,分析其繁殖期的栖息地选择及繁殖对策,探讨这两种鸟类同域共存的机制。
1 研究地区与研究方法
1.1 研究地区
辽宁省东部山区为长白山脉向西南之延伸部分,直到黄、渤海沿岸,海拔多在200 m以上,最高峰为桓仁县境内的老秃顶子,海拔1367.7 m,属于温带大陆性季风气候区,由于受来自东南沿海湿气团的影响,气候温暖湿润,雨热同季,光照和降雨集中,年均温5—8 ℃ ,年平均降雨量800—1000 mm。辽东山区属长白植物区系,森林植被目前主要以天然的次生阔叶林和人工针叶林为主,植物资源丰富多样,主要树种有赤松(Pinusdensiflora)、蒙古栎(Quercusmongolica)、人工红松(Pinuskoraiensis)、核桃楸(Juglansmandshurica)、花曲柳(Fraxinusrhynchophylla)等[16-18]。
1.2 研究方法
2009—2011年的3—7月,累计在研究地随机悬挂山雀式人工巢箱370个,用以招引杂色山雀和大山雀。3a间,共招引到杂色山雀50巢,大山雀65巢。每年4月中旬开始对巢箱进行监测,每7d观察1次,若观察到巢箱底部出现巢材,则每隔1d观察1次,对有卵的巢箱每天观察,记录繁殖情况和以下的繁殖参数:
(1)卵大小 用游标卡尺测量卵长径和短径(精度0.01 mm),用电子秤测量卵重(精度0.01g),卵大小计算公式[19]:V=0. 51 ×L/ mm(卵长径) ×(B/ mm)2(卵短径);
(2)产首枚卵日期 以观察到巢箱中出现第1枚卵为准,如未观察到首枚卵,按每天产1枚卵推测;
(3)在育雏期间,用网捕法对亲鸟进行环志,以观察其是否存在二次繁殖;
(4)孵化期 产下最后1 枚卵到孵出第1 只雏鸟之间的日期;
(5)育雏期 巢内全部雏鸟孵出的日期至最后一只雏鸟出飞的日期;
(6)孵化率 孵出雏鸟的总卵数占繁殖成功巢中总卵数的百分比;
(7)出飞率 出飞雏鸟数占孵出雏鸟数的比例;
(8)繁殖成功率 指繁殖成功的巢数占总巢数的比例,繁殖成功巢是指至少有1 只雏鸟存活到离巢;
(9)繁殖力 按金春日等的方法计算,即繁殖力=平均窝卵数×孵化率×每年繁殖窝数/2[20]。
巢生境选择以巢树为中心,取10 m ×10 m 为样方,对样方中的生态因子进行调查测量,再在巢周围随机选择1个没被利用的人工巢箱作为对照样方。调查项目共19个生态因子,包括:
(1)巢树倾斜角 用罗盘仪测量,单位为度;
(2)巢树高 激光测距仪测量,单位为m;
(3)巢树胸径 用直尺测量,单位为cm;
(4)巢距地高 用卷尺测量,单位为m;
(5)巢口方向 用罗盘仪测量,以正北方向为0°;依顺时针方向,单位为(°);
(6)坡度 用罗盘仪测量,单位为(°);
(7)坡向 用罗盘仪测得,以正北方向为0°;依顺时针方向;
(8)距水源距离 单位为m ;
(9)距路距离 单位为m;
(10)乔木种类 即样方内胸径>4 cm的树木种类;
(11)乔木数量 即样方内胸径>4 cm的树木数量;
(12)乔木盖度 估计值,用百分比表示;
(13)乔木胸径 即样方内胸径>4 cm的平均胸径,单位为cm;
(14)乔木平均高度 单位为m;
(15)灌木种类 样方内高度>30 cm的灌木种类;
(16)灌木盖度 估计值,用百分比表示;
(17)灌木平均高度 单位为m;
(18)草本盖度 估计值,用百分比表示;
(19)植被类型 根据优势乔木的种类,分为阔叶林、针叶林和针阔混交林三类,取值分别为1、2、3。
1.3 数据处理
利用统计软件SPSS(version 17.0)和Excel执行数据统计与分析,利用单个样本的 Kolmogorov-Smirnov Z-检验数据是否呈正态分布,使用经检验符合正态分布的生态因子进行下一步分析。将杂色山雀巢址样方和大山雀巢址样方数据分别与对照样方数据进行T-检验,以比较两种鸟类巢址样方的特异性和各变量差异程度,分析两组数据之间差异的显著水平(<0.05 差异显著,<0.01 差异极显著)。将筛选出的差异显著的变量进行逐步判别分析,找出影响两种鸟类在各自巢址选择过程中的最重要生态因子,并加以比较分析。给出值均为平均值±标准差。
2 结果与分析
2.1 产首枚卵日期及窝卵数
3a间,共招引杂色山雀50巢,大山雀65巢。如图1所示,两种鸟类产首枚卵起始日期大致相同,均为4月中旬,但从种群的产卵期时长上比较的话,大山雀较杂色山雀约长出10 d,杂色山雀最晚于7月上旬产卵,大山雀最晚在7月下旬。两种鸟类均有两次繁殖高峰,大山雀的第1次繁殖高峰为4月中旬至5月上旬,第2次繁殖高峰为5月下旬至6月中旬;杂色山雀的第1次繁殖高峰为4月下旬至5月中旬,第2次繁殖高峰为6月上旬。大山雀第1次繁殖高峰较杂色山雀约提前10d开始,并提前10d结束,第2次繁殖高峰杂色山雀仅持续10d左右,明显短于大山雀(约30d)。两种鸟类的窝卵数在其繁殖期并不是一成不变的,均有下降的趋势,但大山雀窝卵数下降的趋势更为明显,这可能是鸟类根据环境变化而调整其繁殖对策导致的(图中数值为该时期的平均窝卵数)。

图1 巢中首枚卵产出日期与窝卵数比较
2.2 大山雀和杂色山雀繁殖参数比较
大山雀的窝卵数为6—13枚,最常见的为8—11枚卵;杂色山雀的窝卵数为6—9枚,其中产7枚卵的巢数最多(表1)。大山雀与杂色山雀的繁殖参数比较见表2,两者在窝卵数,卵大小,卵重和出飞率上差异极显著(P<0.01),出雏数和出飞数差异显著(P<0.05),孵化率、孵化期、育雏期上差异不显著(P>0.05)。大山雀的窝卵数、出雏数、出飞数和出飞率均大于杂色山雀,而卵大小,卵重则小于杂色山雀。由于受到人为干扰和天敌捕食等原因的影响,大山雀和杂色山雀均有弃巢现象。大山雀的繁殖成功率低于杂色山雀,但繁殖力较高。

表1 繁殖巢数与窝卵数统计

表2 大山雀和杂色山雀的繁殖参数比较
2.3 大山雀的巢址特征
将大山雀巢址样方和对照样方进行Kolmogorov-Smirnov Z检验,将不符合正态分布的巢口方向和坡向两个生态因子不予采用,然后将两组数据进行独立样本T-检验,结果如表3,两者在巢距地高、坡度、乔木数量和乔木平均高度4个变量上存在显著或极显著差异。为找出影响大山雀巢址选择的主要因子,将筛选出的4个存在显著差异的变量进行逐步判别分析,从中得知乔木平均高度、巢距地高和乔木数量是大山雀在巢址选择时的主要关注因子,判别准确率达到84.6%(表4)。
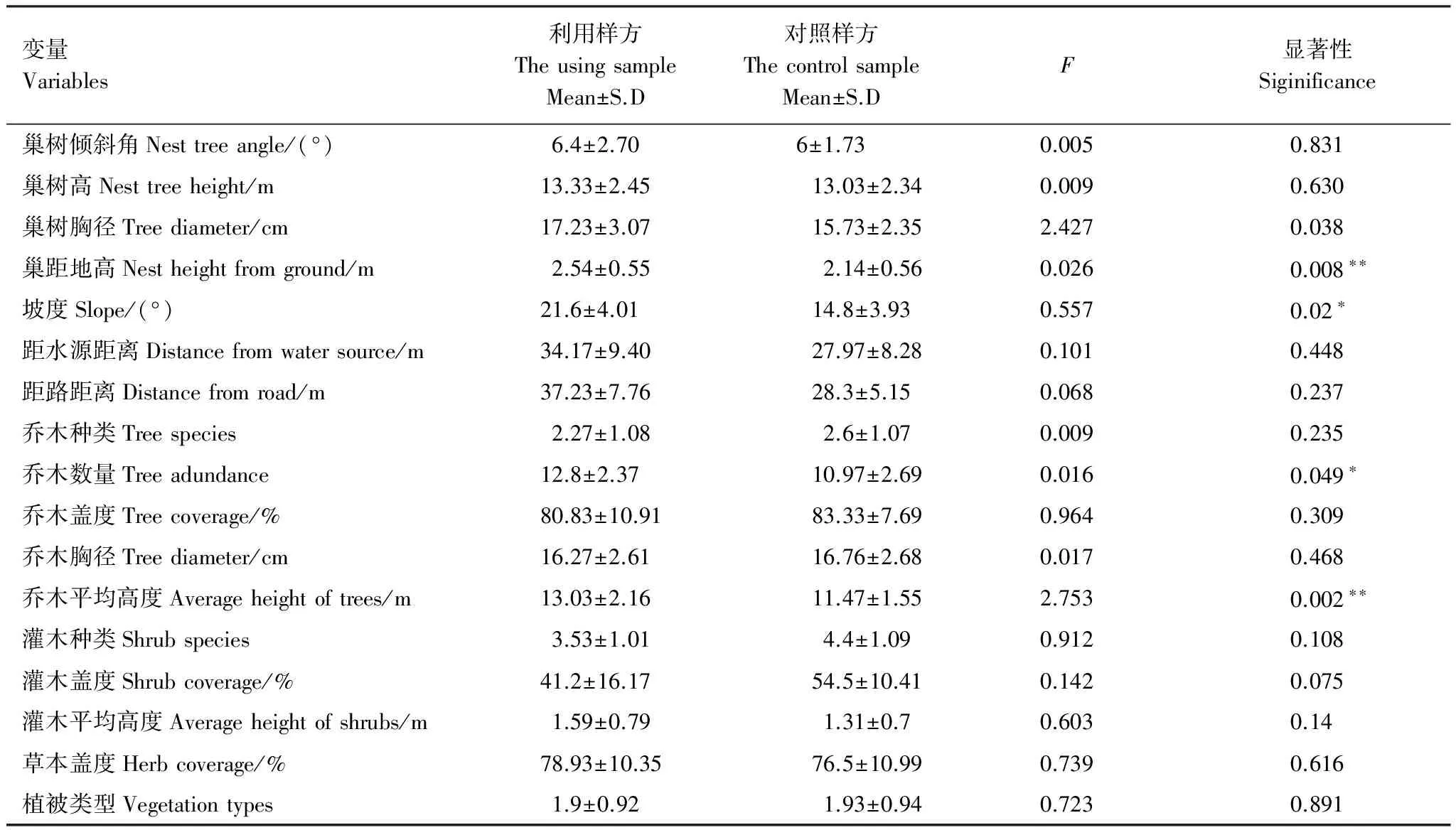
表3 大山雀利用样方和对照样方巢址特征的比较(n=30)
差异显著*P<0.05,差异极显著**P<0.01
2.4 杂色山雀的巢址特征
将杂色山雀巢址样方和对照样方进行Kolmogorov-Smirnov Z检验,将不符合正态分布的巢口方向和坡向两个生态因子不予采用,然后将两组数据进行独立样本T-检验,结果如表5,两者在距水源距离、距路距离、乔木平均高度、灌木平均高度和植被类型5个变量上存在显著或极显著差异(表5)。为找出影响杂色山雀巢址选择的主要因子,将筛选出的5个存在显著差异的变量进行逐步判别分析,从中得知乔木平均高度、植被类型和灌木平均高度是杂色山雀在巢址选择时的主要关注因子,判别准确率达到85.9%(表6)。
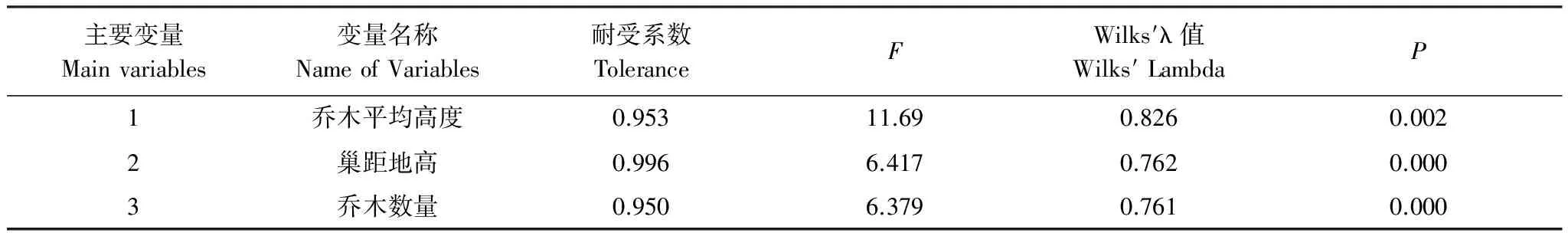
表4 大山雀巢址利用样方与对照样方逐步判别分析

表5 杂色山雀利用样方和对照样方巢址特征的比较(n=24)
差异显著*P<0.05,差异极显著**P<0.01

表6 杂色山雀巢址利用样方与对照样方逐步判别分析
3 讨论
适合度和权衡是生活史对策的两个重要概念。适合度是衡量一个个体存活和繁殖成功机会的一种尺度。权衡是描述动物牺牲一方面的利益,期望在另一方面得到回报的行为活动。生活史对策是在生殖和存活这两个对立需求之间,把有限的资源进行合理地分配[21]。野外研究发现,动物能适时地调整其繁殖对策,以使个体的适合度最大化[22]。
动物的繁殖在每个生命周期中一般是受到时间限制的,繁殖期的时间限制与社群压力的存在决定了生境选择的时间制约性[23],动物繁殖成功的可能性会随着时间而递减,短暂的繁殖季节要求动物必须迅速做出选择决定[24]。在辽宁省东部山区,鸟类适宜的繁殖时间大约为4月上旬至7月下旬,种间和种内竞争十分激烈,大山雀与杂色山雀均为次级洞巢鸟类,人工巢箱的大量悬挂降低了这两种鸟类对洞巢的竞争,但是个体进行生境选择的越晚,可选择的生境就越恶劣,适宜的生境被抢先占据,那些后来者就必须在质量较差的生境中存活,随着时间的推迟,食物等资源逐渐减少,鸟类自身的繁殖成功率会有所下降[25]。大山雀的两次繁殖高峰均较杂色山雀提前,有利于使其在种间竞争中处于优势地位,优先占据那些环境质量较好的巢址,增加繁殖成功的机会。而且大山雀的第2次繁殖高峰较杂色山雀持续时间更长,这说明大山雀可能比杂色山雀更能充分地利用有限的繁殖季节繁衍出更多的后代。另外,本文研究还发现,大山雀和杂色山雀均出现二次繁殖现象,即一对繁殖鸟在一个繁殖季节先后繁殖两次。二次繁殖需要成鸟投入更多的能量,但能够增加鸟类的年繁殖生产力[26]。两种鸟类的窝卵数都有逐渐减少的趋势,说明两者为了适应环境的变化,通过繁殖代价与繁殖成效间的权衡,均采用了消减窝卵数的对策来提高繁殖成功率,提高自身的年繁殖生产力。
MacArthurt将生态对策分为r选择对策和k选择对策,Piankat发展了r/k选择理论,认为在r/k之间有许多过渡类型,同r选择比较可能更接近于k选择,同k选择比较可能更接近于r选择,即动物的生殖对策是r-k选择的连续统一体(r-k continuum)[27-28]。根据种群进化对策理论[29],在稳定、均质环境中,自然选择应偏向低繁殖率和高竞争力的k-选择对策的物种进化;在季节性的可变环境中,则偏向高繁殖率和短寿的r-选择对策的物种进化[30],繁殖对策的选择对鸟类自身的存活和种群的繁衍具有重要意义。程成通过对人工巢箱中大山雀和褐头山雀(Parusmontanus)的繁殖比较研究认为,大山雀可能采取高产卵数、低存活率的繁殖对策,而褐头山雀的繁殖策略则可能为低产卵数、高存活率[4]。本文对大山雀和杂色山雀的繁殖参数比较研究表明,两种鸟类均属于k-选择对策,但大山雀相对接近r 端。与杂色山雀相比:大山雀的窝卵数多,孵化率低,育雏期短,出飞率高,繁殖成功率低,繁殖力高。高窝卵数增加了大山雀繁殖成功的难度,要求其繁殖投入更多,未受精卵较多造成了大山雀孵化率较低,而有时受精卵还未孵化,雌鸟即进入了育雏期,不再对剩余的卵进行孵化,大山雀的体型小于杂色山雀,使得雏鸟用相对少的能量就可以达到出飞条件,较短的育雏期也降低了雏鸟被天敌捕食的概率,提高了大山雀雏鸟的出飞率;大山雀多在距路30m以上的巢箱筑巢,而且隐蔽条件较好,但是大山雀十分机警,在产卵期和孵化前期遇到惊扰极容易选择弃巢,这也是造成大山雀繁殖成功率低于杂色山雀的主要原因。与之相反,低窝卵数提高了杂色山雀的孵化率,但育雏期的延长增加了雏鸟被天敌捕食的机会,使雏鸟出飞率有所下降。繁殖力是种群生长的基础,大山雀的繁殖力为3.36,高于杂色山雀的2.93,高窝卵数、较高的孵化率以及迅速展开的2次繁殖有益于大山雀种群数量的提高。
不同资源在空间上的不均匀分布,使得动物必须在各种资源的空间配置中进行权衡[31-32]。在繁殖季节,鸟类对空间选择首要考虑的就是巢址选择。Cody指出,鸟类的巢址选择是一种优化生境选择,鸟类总是把巢址建立在最利于其繁殖的地方,但鸟类的巢址选择不可能达到十全十美,因为除了物质资源的限制外,还存在着竞争和干扰等因素的影响[33]。适宜的巢址能将同类的干扰作用、天敌捕食和其它不良因子的影响降到最低水平从而提高繁殖成功率,巢址选择是鸟类在长期进化过程中形成的一种繁殖与防御对策[34],许多鸟类都倾向于选择那些能使其繁殖成效最大而存活代价最小的营巢生境[35]。通过与对照样方的比较可以看出,大山雀多筑巢于乔木高大且数量较多、巢距地较高、有一定坡度的森林中,杂色山雀多选择在距水、距路较近,乔木、灌木生长茂盛的针阔混交林边缘筑巢。从逐步判别分析的结果看,乔木平均高度是两种鸟类在进行巢址选择时的首要关注因子。很多森林中筑巢的鸟类多选择周围乔木高大的巢址,如啄木鸟 (Melanerpeslewis)[36]、赤肩鵟(Buteolineatus)[37]。乔木的高度反映的是林龄,林龄长的森林中存在更多的天然巢址,物种多样性也较为丰富,食物更充足,可以使鸟类不用远行就能找到足够的昆虫喂养雏鸟,减少成鸟在育雏期的能量支出。大山雀的巢址距地面较高,周围乔木生长密集,也就增加了乔木的密度和盖度,这反映了鸟类对巢址安全性能的要求,巢址距地较高可以降低雏鸟被地面天敌(如蛇、鼠)发现和捕食的概率(野外研究发现,蛇捕食是山雀类雏鸟死亡的主要原因),许多小型鸟类均比较注重巢址的安全性能,如加拿大威森莺(Wilsoniacanadensis)[38];杂色山雀则对植被类型有所选择,不同植被类型的森林中乔木、灌木的种类和数量会有所差异,针阔混交林中灌木生长茂盛,其中分布的昆虫种类更加多样,杂色山雀偏爱在针阔混交林中筑巢,可能是出于食物方面的考虑。
本研究认为,作为在辽东山区同域分布的两种小型森林洞巢鸟类,生态位的部分分离是大山雀和杂色山雀能够长期共存的基础。两者都是在繁殖期到来即开始繁殖,为了提高年繁殖生产力,均利用短暂的繁殖季节进行第2次繁殖,采用消减窝卵数的对策来适应环境质量的下降,已达到提高繁殖成功率的目的。两种鸟类的繁殖对策都为k-选择,但大山雀略偏向r端,相对而言,大山雀倾向高窝卵数、低繁殖成功率的繁殖策略;杂色山雀倾向低窝卵数、高繁殖成功率的繁殖策略。两者在巢址选择上虽各有偏好,但都选择在林龄较长的森林中筑巢,大山雀较注重巢址的安全性能,巢址距地面较高,周围乔木密集;杂色山雀较为注重植被类型的选择,巢址多选择在植被多样、灌木生长茂盛的针阔混交林中,这可能与杂色山雀的食物构成有关。本研究的不足之处是缺乏两种鸟类在食性方面的比较,食性作为鸟类繁殖策略的重要组成部分,将在今后的工作中加以补充。
[1] Li B, Yang C, Lin P. Ecology. Beijing: Higher Education Press, 2000.
[2] Sun R Y. Principles of Animal Ecology. 3rd ed. Beijing: Beijing Normal University Press, 2001: 71-71.
[3] Yin B F, Wei W H, Zhang Y M, Cao Y F, Wang J L. Reproductive investment and reproductive success of small mammals. Acta Theriologica Sinica, 2003, 23(3): 259-265.
[4] Cheng C, Liang W, Zhang Z H. Reproductive strategies of great tit and willow tit in artificial nest boxes in Xiaolongmen forest park of Beijing. Chinese Journal of Ecology, 2011, 30(7): 1575-1578.
[5] Ghalambor C K, Martin T E. Parental investment strategies in two species of nuthatch vary with stage-specific predation risk and reproductive effort. Animal Behaviour, 2000, 60(2): 263-267.
[6] Zhao L. Behavioral responses of two species passerine to predation risk during breeding period. Zoological Research, 2005, 26(2): 113-117.
[7] Minot E O. Effects of interspecific competition for food in breeding Blue and Great tits. Journal of Animal Ecology, 1981, 50(2): 375-385.
[8] Dhondt A A. Effects of competition on great and blue tit reproduction: intensity and importance in relation to habitat quality. Journal of Animal Ecology, 2010, 79(1): 257-265.
[9] Zhou F, Fang H L. On the interspecific niche relationship between two species of wren warbler. Zoological Research, 2000, 21(1): 52-57.
[10] Park Y S, Lee W S, Rhim S J. Influence of forest road on breeding of tits in artificial nest boxes. Journal of Forestry Research, 2005, 16(4): 301-302.
[11] Cui P, Kang M J, Deng W H. Foraging habitat selection by sympatric temminck′s tragopan and blood pheasant during breeding season in southwestern China. Biodiversity Science, 2008, 16(2): 143-149.
[12] Wang W K, Zhou C Q, Long S, Yang W Z, Liu Y D, Hu J C. Spartial niche and interspecific relationships of ardeidae birds in Nanchong Taihe little egret nature reserve. Sichuan Journal of Zoology, 2008, 27(2): 178-182.
[13] Zhao L, Zhang X A. Nest-site selection and competition coexistence of horned larks and small skylarks. Zoological Research, 2004, 25(3): 198-204.
[14] Zhao Z J. Chinese Avifauna (Passeriformes). Changchun: Jilin Publing House of Science and Technology, 2001: 688-690.
[15] Zheng G M. A Checklist on the Classification and Distribution of the Birds of China. Beijing: Science Press, 2005: 328-329.
[16] Liu M Y, Qiu Y J, Li Z W. Birds Survey Report in Liaoning Province. Shenyang: Liaoning University Press, 1986: 60-63.
[17] Sheng X J, Wang S G, Guan D X, Jin C J, Wang A Z, Wu J B, Yuan F H. Canopy interception in larch plantations: measurement and modeling in eastern Liaoning mountainous region. Chinese Journal of Applied Ecology, 2010, 21(12): 3021-3028.
[18] Jiao X L, Zhu J J, Yan Q L. Composition and seasonal dynamics of macro-and meso-soil fauna in secondary forest ecosystems of Liaodong montane area, China. Acta Ecologica Sinica, 2009, 29(5): 2631-2638.
[19] Yuan S C, Zhang K Q, Li T. A comparison study on the population ecology of Paridae birds in Changbai Mountain. Journal of Jilin Forestry Institute, 1994, 10(3): 141-146.
[20] Jin C R, Wang S, Wang D M, Li H Y, Qu Z C, Ding F. Reproductive Success ofParusvariusin natural condition and nest-box. Chinese Journal of Zoology, 2007, 42(3): 28-33.
[21] Gao W. Studies on Birds and Their Ecology in Northeast China. Beijing: Science Press, 2006: 257-257.
[22] Stearns S C. The Evolution of Life Histories. Oxford: Oxford University Press, 1992.
[23] Zhang M H, Li Y K. The temporal and spatial scales in animal habitat selection research. Acta Theriologica Sinica, 2005, 25(4): 395-401.
[24] Orians G H, Wittenberge J F. Spatial and temporal scales in habitat selection. The American Naturalist, 1991, 137(S1): S29-S49.
[25] Pimm S L, Rosenzweig M L, Mitchell W. Competition and food selection: field tests of a theory. Ecology, 1985, 66(3): 798-807.
[26] Qin B, Jiang Y L, Li S, Zhang L S, Wang H T, Gao W. Comparison of reproductive parameters between the first and second broods in Great Tit. Journal of Northeast Normal University: Natural Science Edition, 2010, 42(2): 101-104.
[27] MacArthur R H, Wilson E O. The Theory of Island Biogeography. New Jersey: Princeton University, 1967.
[28] Nie H Y, Liu J K, Su J P, Zhang Y M, Zhang H H. Progress in the study of animal life history evolution. Acta Ecologica Sinica, 2007, 27(10): 4267-4277.
[29] MacArthur R H. Geographical Ecology. Princeton: Princeton University Press, 1972.
[30] Wang C. Evolutionary Ecology. Beijing: Higher Education Press, 2008: 192-195.
[31] Brown J S. Patch use as an indicator of habitat preference, predation risk, and competition. Behavioral Ecology and Sociobiology, 1988, 22(1): 37-47.
[32] Lima S L, Dill L M. Behavioral decisions made under the risk of predation: a review and prospectus. Canadian Journal of Zoology, 1990, 68(4): 619-640.
[33] Cody M L. Habitat Selection in Birds. Orlando: Academic Press, 1987.
[34] Cody M L. Habitat selection in birds: the roles of vegetation structure, competitors and productivity. Bioscience, 1981, 31(2): 107-113.
[35] You Y Q, Zhou C Q, Hu J C. Nest-building and breeding behavior of Tiger Shrike (Laniustigrinus). Sichuan Journal of Zoology, 2007, 26(1): 26-31.
[36] Newlon K R, Saab V A. Nest-Site selection and nest survival of Lewis′s woodpecker in aspen riparian woodlands. The Condor, 2011, 113(1): 183-193.
[37] King J C, Dubay S A, Woodford J E. Distribution and nest site selection of red-shouldered hawks (Buteolineatus) in forests of northeastern Wisconsin (USA). Forest Ecology and Management, 2011, 261(1): 169-177.
[38] Goodnow M L, Reitsma L R. Nest-site selection in the Canada Warbler (Wilsoniacanadensis) in central New Hampshire. Canadian Journal of Zoology, 2011, 89(6): 1172-1177.
参考文献:
[1] 李博, 杨持, 林鹏. 生态学. 北京: 高等教育出版社, 2000.
[2] 孙儒泳. 动物生态学原理 (第三版). 北京: 北京师范大学出版社, 2001: 71-71.
[3] 殷宝法, 魏万红, 张堰铭, 曹伊凡, 王金龙. 小型哺乳动物的繁殖投入与繁殖成功率. 兽类学报, 2003, 23(3): 259-265.
[4] 程成, 梁伟, 张子慧. 人工巢箱中大山雀和褐头山雀的繁殖比较. 生态学杂志, 2011, 30(7): 1575-1578.
[6] 赵亮. 繁殖期两种百灵科鸟类对捕食风险的行为响应. 动物学研究, 2005, 26(2): 113-117.
[9] 周放, 房慧伶. 两种鹪莺的种间生态位关系研究. 动物学研究, 2000, 21(1): 52-57.
[11] 崔鹏, 康明江, 邓文洪. 繁殖季节同域分布的红腹角雉和血雉的觅食生境选择. 生物多样性, 2008, 16(2): 143-149.
[12] 王维奎, 周材权, 龙帅, 杨文仲, 刘延德, 胡锦矗. 四川南充太和鹭科鸟类群落空间生态位和种间关系. 四川动物, 2008, 27(2): 178-182.
[13] 赵亮, 张晓爱. 角百灵和小云雀的巢址选择与竞争共存. 动物学研究, 2004, 25(3): 198-204.
[14] 赵正阶. 中国鸟类志(雀形目). 长春: 吉林科学技术出版社, 2001: 688-690.
[15] 郑光美. 中国鸟类分类与分布名录. 北京: 科学出版社, 2005: 328-329.
[16] 刘明玉, 邱英杰, 李壮威. 辽宁鸟类考查报告. 沈阳: 辽宁大学出版社, 1986: 60-63.
[17] 盛雪娇, 王曙光, 关德新, 金昌杰, 王安志, 吴家兵, 袁凤辉. 辽宁东部山区落叶松人工林林冠降雨截留观测及模拟. 应用生态学报, 2010, 21(12): 3021-3028.
[18] 焦向丽, 朱教君, 闫巧玲. 辽东山区次生林生态系统大、中型土壤动物组成与季节动态. 生态学报, 2009, 29(5): 2631-2638.
[19] 袁守城, 张克勤, 李彤. 长白山山雀科鸟类种群生态的比较研究. 吉林林学院学报, 1994, 10(3): 141-146.
[20] 金春日, 王爽, 万冬梅, 李昊晔, 曲再春, 丁峰. 杂色山雀的繁殖成功率. 动物学杂志, 2007, 42(3): 28-33.
[21] 高玮. 中国东北地区鸟类及其生态学研究. 北京: 科学出版社, 2006: 257-257.
[23] 张明海, 李言阔. 动物生境选择研究中的时空尺度. 兽类学报, 2005, 25(4): 395-401.
[26] 秦博, 姜云垒, 李时, 张立世, 王海涛, 高玮. 大山雀一次和二次繁殖参数比较. 东北师大学报: 自然科学版, 2010, 42(2): 101-104.
[28] 聂海燕, 刘季科, 苏建平, 张堰铭, 张洪海. 动物生活史进化理论研究进展. 生态学报, 2007, 27(10): 4267-4277.
[30] 王崇云. 进化生态学. 北京: 高等教育出版社, 2008: 192-195.
[35] 游余群, 周材权, 胡锦矗. 虎纹伯劳的巢生境选择与繁殖行为. 四川动物, 2007, 26(1): 26-31.
