青藏高原高寒草甸退化与人工恢复过程中植物群落的繁殖适应对策
2013-12-08李媛媛董世魁李小艳王学霞
李媛媛,董世魁,朱 磊, 温 璐, 李小艳,王学霞
(北京师范大学环境学院,北京 100875 )
青藏高原高寒草甸退化与人工恢复过程中植物群落的繁殖适应对策
李媛媛,董世魁*,朱 磊, 温 璐, 李小艳,王学霞
(北京师范大学环境学院,北京 100875 )
以三江源区不同退化程度高寒草甸和不同恢复年限人工草地作为研究对象,通过野外调查与采样、实验室分析,探究了高寒地区退化天然草地与人工恢复草地的植被群落繁殖构件数量变化。结果表明:在群落水平上,天然草地退化和人工草地建植会对植物繁殖构件的数量和生物量产生影响。随着天然草地退化程度的增加,营养枝数量和生物量则明显下降,而繁殖枝的数量和生物量明显升高(P<0.05);随着人工草地恢复年限的增加,营养枝的数量和生物量逐渐增加,而繁殖枝的数量和生物量则逐渐降低(P<0.05);随着恢复年限的增加,人工草地繁殖构件的变化逐渐接近未退化天然草地。在功能群水平上,植物繁殖构件数量亦随草地退化程度和人工恢复年限而变化。随着恢复年限的增加,禾本科、莎草科、杂类草的营养枝数量和生物量均呈现显著增加(P<0.05),而繁殖枝数量和生物量则显著下降,禾本科的繁殖构件数量远远大于莎草科和杂类草;随着退化程度的增加,三大功能群的营养枝枝数和生物量显著增加(P<0.05),而繁殖枝则呈现相反的趋势。实证了草地退化和人工恢复改变植物群落繁殖分配对策的科学假设,为高寒草地植被恢复重建技术的发展和更新提供理论支撑。
青藏高原;退化草地;人工草地;植物群落;繁殖对策
三江源区位于青藏高原腹地,是青藏高原的主体部分,同时也是我国的最重要的水源地,素有“中华水塔”之称[1]。其独特的自然状况和地理位置决定了该区高寒草地特有的生态功能和生产功能,对于区域水资源、生物资源、草畜平衡、全球气候变化等方面起到了极其重要的作用[2]。三江源地区草地以高寒草甸为主,然而,近年来,由于气候变化和人类活动的影响,三江源地区高寒草甸生态系统的功能和结构极度受损,呈现了逐步恶化的趋势,严重影响了该地以及周边地区社会、经济、环境的可持续发展[3- 5]。针对三江源地区草地退化的问题,不少学者提出了一系列的恢复措施,包括围栏封育、建立人工草地、草地补播、鼠虫害及毒杂草防除等措施[6]。这些措施为高寒草地生态环境的建设与保护提供了技术支撑。相对而言,退化高寒草地植被恢复重建的基础理论研究相对薄弱,一定程度上限制了恢复重建技术措施的更新和发展[7]。
从理论上讲,退化草地的成功恢复主要依赖于两个方面:植被繁殖更新潜力和土壤养分供给能力。就前者而言,草地群落中可利用繁殖体(种子和母株无形繁殖幼苗)提供了退化草地得以恢复的内在条件及潜在动力[8],是退化草地生态恢复的主要研究内容,近年来被许多科学研究工作者所关注[9- 11]。在高寒生境中大部分植物采用了以营养繁殖为主、有性繁殖为辅的繁殖策略[12]。有性繁殖可以影响植物对于当地环境的适应性以及基因进化的过程,而无性繁殖有利于植物的传粉过程,提高植物对于环境的适合度[13]。草地退化造成了土壤养分和物理特性的差异,可能会影响克隆植物某些特性的表达,从而造成草地繁殖对策的改变[14]。目前,对青藏高原地区草地植被繁殖动力学的研究主要集中在三个方面:一是以生态适应性理论为基础,分析人工或者自然调控状态下高寒草地主要植物种群的繁殖对策[15- 16];二是以阈值理论为基础,探讨极端生境条件或退化状态对高寒草地植物繁殖模式的塑造作用[17- 18];三是以干扰理论为基础,研究不同放牧压力对高寒草地植物繁殖模式的选择作用[19- 20]。这些研究从不同方面深入分析了矮嵩草、垂穗披碱草等几种主要高寒草地植物的繁殖模式,揭示了这些高寒植物适应极端环境和退化生境、抵御放牧干扰的繁殖对策。但是,仍缺乏群落尺度植物繁殖生态机制研究,尚不能全面反映高寒草地植物群落恢复演替的繁殖动力学过程。
因此,本文基于草地退化和人工恢复改变植物群落繁殖分配对策的科学假设,以三江源高寒草地为研究对象,分析不同退化程度天然草地和不同恢复年限人工草地的植物群落繁殖对策,以期回答如下3个科学问题:(1)草地退化和人工恢复是否会改变植物群落的繁殖策略?(2)在草地退化和人工恢复序列上植物群落的繁殖分配是如何变化的?(3)在草地退化和人工恢复序列上植物群落的繁殖构件数量是如何变化的?这些科学问题的准确解答将促进高寒草地植被生态恢复理论的纵深发展,从而为高寒草地植被恢复重建技术的发展和更新提供理论支撑。
1 材料与方法
1.1 研究区概况
本研究于2009—2010年在青海省果洛藏族自治州玛沁县大武镇(N32°31′—35°37′,E96°54′—101°51′)进行。该区位于我国著名的“江河源区”,以巴颜喀拉山为分水岭,形成长江、黄河两大水系。该区境内海拔高度多在4000 m以上,光照时数月均2000 h左右,年均气温-1.3℃,最冷月1月均温-12.4℃,最热月7月均温9.8℃,年均降水486.9—666.5 mm,多集中在6—9月,年蒸发量为1119.1 mm,属高寒半湿润型气候[21]。自然植被以高山(寒)草甸为主,土壤为高山(寒)草甸土,无绝对无霜期,植物生长期达120—140 d,是典型的高寒地区。该区天然草地在气候变化和过度放牧等自然和人为因素的影响下,出现严重退化的现象,部分地区已成为“黑土滩”,即以嵩草属植物为建群种的草地植被严重退化后已被铁棒锤(Aconitumpendulum)、甘肃马先蒿(Pediculariskansuensis)、黄帚橐吾(Ligulariavirgaurea)、细叶亚菊(Ajaniatenuifolia)等代替,优良牧草盖度不足10%,基本不具备牧用价值。夏季植被总盖度可达70%,但多以阔叶毒杂草为主,在冬、春季节地面基本处于裸露状态。
2 研究方法
本文根据马玉寿[22]提出的高寒草地退化等级划分方法,根据植被盖度、高度、生物量等指标分别在研究区选择未退化、中度退化、重度退化和极度退化高寒草甸作为不同退化程度草地的研究对象,每类草地随机选择3个斑块(面积500 m2—1hm2)作为重复样地。同时,选取建植5a、7a、9a的垂穗披碱草人工草地作为不同恢复年限草地的研究对象,每类草地随机选择3个实验小区作为重复样地。在草地植物旺盛生长期(7—8月),采用董鸣[23]介绍的方法调查植物群落结构和数量特征,得出草地群落的物种组成及建群植物(表1)。然后,采用周兴民[24]介绍的方法,测定草甸植被主要种群繁殖对策参数,具体做法为:在各个样地上布设3个50 cm×50 cm样方,将植物群落建群种(表1)的繁殖枝和营养枝进行计数并齐地剪取,然后进行分装、烘干、称重,同时记录繁殖枝上的小穗及种子数目;对于根茎、根蘖、直根等地下营养繁殖器官,在各样方内随机选择10个采样点,每个样方用土钻(直径3.5 cm)分层(0—4 cm、4—10 cm、10—20 cm)共取30钻土壤(每层10钻),采用漂洗法将根洗净(将土样通过0.45 mm的尼龙袋,土壤中的根状物将残留在尼龙袋中),将0—4 cm土层中的根的根茎、根蘖、直根等营养繁殖体数目进行计数及称重(鲜重及干重),其原因在于根据前人[25]研究该区0—4cm的土层富含根系但易被剥蚀(水蚀或风蚀);其余土层的样品仅烘干、称重,以计算总根量测重。
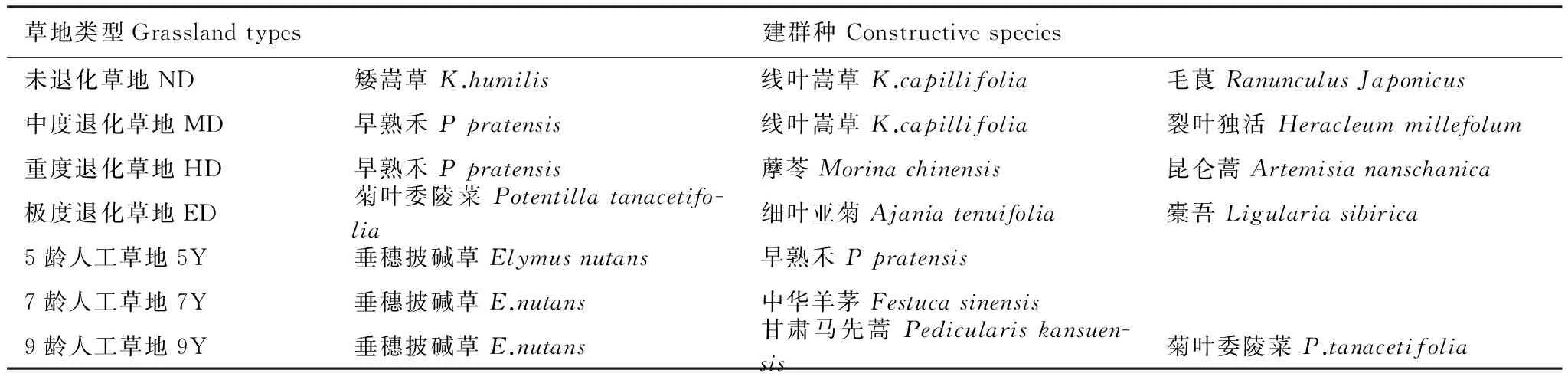
表1 天然草地和人工草地的建群种组成
5Y: five years′ restoration artificial grassland,7Y: seven years′ restoration artificial grassland, 9Y: nine years′ restoration artificial grassland, ND: non-degraded grassland, MD: moderately degraded grassland, HD: heavily degraded grassland, ED: extremely degraded grassland
2.1 数据处理与统计分析
将所有样方调查数据求算平均值,运动SPSS16.0 软件对其进行方差分析,利用LSD法对其进行差异性检验,比较不同处理下草地植物群落生殖枝、营养枝的数量和重量差异。
3 结果分析
3.1 不同处理下营养枝数量和生物量的变化
如图1所示,随着恢复年限的增加,地上营养枝数量逐渐增加,其中9龄人工草地的营养枝枝数远远高于5龄和7龄的营养枝枝数(P<0.05);随着退化程度的增加,营养枝数量也呈现降低的趋势。总体上,天然草地的营养枝指数低于人工恢复草地的营养枝指数。营养枝生物量的变化趋势与营养枝枝数的变化趋势基本一致(图1),天然草地与人工草地的营养枝的生物量无显著差异,但不同生长年限的人工草地营养枝生物量存在显著性差异(P<0.05)。其中,5龄人工草地营养枝生物量与极度退化草地的生物量相近 (P>0.05),7龄人工草地营养枝生物量与重度退化草地无显著性差异(P>0.05)。

图1 不同处理草地类型营养枝枝数和营养枝生物量的变化(平均值±标准方差)Fig.1 Changes about the numbers and biomass of vegetative branches in different treatment grasslands5Y: 5龄人工恢复草,7Y: 7龄人工恢复草地,9Y: 9龄人工恢复草地,ND: 未退化草地,MD: 中度退化草地,HD: 重度退化草地,ED: 极度退化草地5Y: five years′ restoration artificial grassland,7Y: seven years′ restoration artificial grassland, 9Y: nine years′ restoration artificial grassland, ND: non-degraded grassland, MD: moderately degraded grassland, HD: heavily degraded grassland, ED: extremely degraded grassland不同柱间的相同字母表示无显著性差异
3.2 不同处理下繁殖枝数量和生物量的变化
繁殖枝数量和生物量与营养枝呈现相反的趋势(图2)。繁殖枝枝数随着恢复年限的增加逐渐降低(P<0.05),且5龄人工草地的繁殖枝数量远远大于另外两类人工草地(P<0.01). 随着退化程度的增加,草地群落的繁殖枝数目逐渐升高。总体上,退化草地的繁殖枝生物量小于人工草地(图2)。随着恢复年限的增加,人工草地群落的繁殖枝生物量呈现下降的趋势;随着退化程度的增加,天然草地群落的繁殖枝生物量也呈下降趋势。
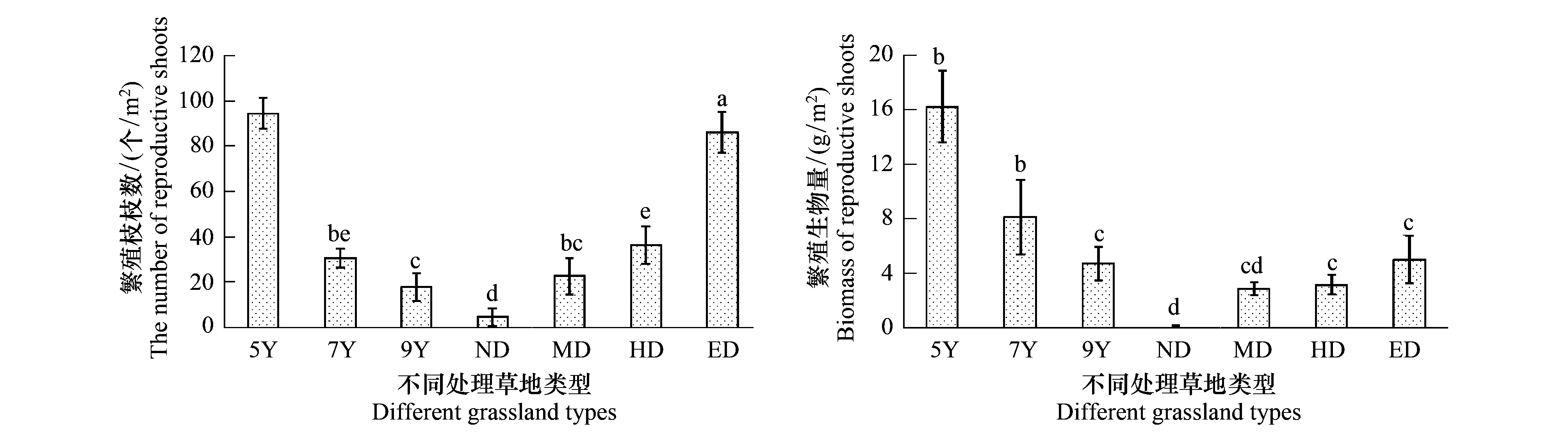
图2 不同处理草地类型繁殖枝枝数和生物量的变化(平均值±标准方差)Fig.2 Changes about the numbers and biomass of reproductive branches in different treatment grasslands (mean ±SD)
3.3 不同处理下群落无性繁殖总构件数量和生物量的变化
将地上营养枝和地下繁殖构件作为群落无性繁殖总构件进行分析,得出其变化趋势与营养枝基本一致(图3)。随着恢复年限的增加,人工草地植物群落无性繁殖总构件呈现上升的趋势(P<0.05)。随着退化程度的加剧,天然草地植物群落无性繁殖总构件呈现下降的趋势(P<0.05)。尽管人工恢复草地无性繁殖总构件数量小于未退化草地,但差距随着恢复年限的增加而减少(图3)。总体上,人工恢复草地无性繁殖总构件的生物量小于天然草地,但随着恢复年限的增加,这一差距逐渐减小(图3);未退化草地无性繁殖总构件的生物量显著大于其他几类草地(P<0.05)。

图3 不同处理草地类型无性繁殖总构件数量和生物量的变化(平均值±标准方差)Fig.3 Changes about the numbers and biomass of asexual reproduction component in different treatment grasslands (mean ±SD)
3.4 不同处理下群落繁殖分配的变化
从繁殖分配的角度看,未退化天然草地的有性繁殖分配几乎为零(图4)。人工草地有性繁殖的比例随着恢复年限的增加显著降低(P<0.05),而天然草地有性繁殖的比例并不随退化程度发生显著变化(P>0.05)。总体上,人工恢复草地的有性繁殖比例大于退化天然草地(P<0.05)。
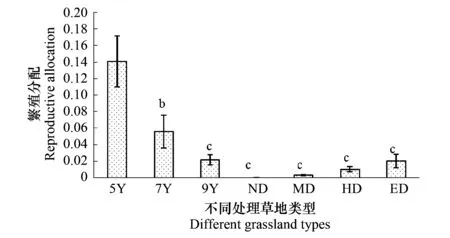
图4 不同处理草地类型繁殖分配的变化(平均值±标准方差)Fig.4 Changes about the reproductive allocation in different treatment grasslands
3.5 不同处理下主要植物类群的繁殖构件的变化
随着人工草地恢复年限的增加,禾本科植物类群营养枝数及生物量都逐渐增加,而繁殖枝枝数及生物量则呈现相反的趋势(图5),并且营养枝的数量和生物量远远大于繁殖枝枝数及生物量(P<0.05)。随着天然草地退化程度的加剧,禾本科植物类群的营养枝枝数逐渐减少,而繁殖枝枝数和生物量则呈现相反的趋势;除重度退化和极度退化草地外,其他草地的繁殖枝枝数和生物量都达到了极显著差异(P<0.01)。
对于莎草科植物而言,营养枝枝数随着天然草地退化程度的加剧而减少(P<0.05),而繁殖枝枝数随着天然草地退化程度的加剧而增加。在未退化天然草地中,该类群的繁殖枝枝数及生物量都处在最低水平。
对杂类草植物而言,人工草地和天然草地上的营养枝和繁殖枝枝数及生物量的变化趋势与禾本科类群基本一致。

图5 不同处理草地类型禾本科营养枝、繁殖枝数量和生物量的变化(平均值±标准方差)Fig.5 Changes about the numbers and biomass of vegetative and reproductive branches of grass in different treatment grasslands (mean±SD)
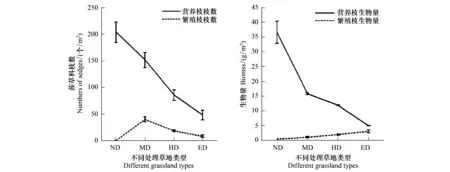
图6 不同处理草地类型莎草科营养枝、繁殖枝数量和生物量的变化(平均值±标准方差)Fig.6 Changes about the numbers and biomass of vegetative and reproductive branches of sedges in different treatment grasslands (mean±SD)
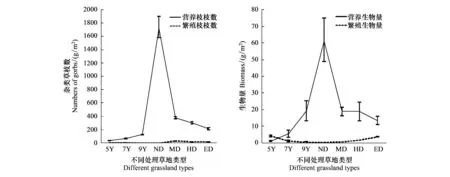
图7 不同处理草地类型杂类草禾本营养枝、繁殖枝数量和生物量的变化(平均值±标准方差)Fig.7 Changes about the numbers and biomass of vegetative and reproductive branches of forbs in different treatment grasslands (mean±SD)
3 讨论与结论
3.1 退化和恢复草地的植物繁殖适应对策
植物的繁殖对策是指植物在其生活史过程中,通过最佳的资源分配格局,以其特有繁殖属性的适应过程,提高自身适合度的组织过程[26]。研究植物在不同环境中的繁殖对策能够反映出植物对环境的适应能力和在该环境中的生殖潜能。在草地群落中,许多植物物种是具有匍匐茎或者根茎的多年生草本植物,它们多以无性繁殖或称为克隆繁殖进行个体数量的补充和增殖,所以,种群数量的变化主要体现在构件数量而并非源株数量的变化上[20]。且营养繁殖的主要优势为子株的死亡率较低[27],Schimid 的研究表明,此种增值方式对群落中特定种多度的影响要远大于种子扩散的作用[28]。本文的研究结果表明,从繁殖分配角度人工草地有性繁殖分配随着恢复年限的增加显著降低,而天然草地随着退化程度的增加显著增加。这可能因为在无干扰的环境中,植物长期处于强烈竞争状态,在这种竞争下,无性系的分株通常比种子形成的幼苗更加容易成活[29],并且青藏高原特殊的严酷生长环境也限制了大量种子的萌发。通常情况下,草原生态系统受人和牲畜干扰比较大,再加上降雨季节性不平衡,使得草原生态系统形成了自己独特的繁殖方式。尽管草原上大部分植物为克隆植物,并且这些植物几乎都能产生种子,但有性后代较少,很大程度上是跟春季干旱的环境有关[30]。相反,高寒环境中植物营养繁殖通常适合度较高,具有进化的优势,有利于提高个体的生存和竞争力,从而有利于种群的迅速扩散和传播[31]。
3.2 群落水平上的植物繁殖分配对策
植物的繁殖特征受环境因子的影响强烈,随环境干扰因子及强度的变化,草地植物有很大的形态变化[32- 33],例如群落结构的变化,土壤养分有效性和下垫面微气候特征变化等,都会对物种的生态适应策略产生一定程度的影响[33]。本文对营养枝和繁殖枝数量与生物量的研究结果表明表明,随着退化程度的加剧,营养枝数和生物量下降而繁殖枝数和生物量增加。说明草地退化不有利于营养枝生物量的积累,而有利于生殖枝生物量的积累。这可能是由于健康草地植物个体密度加大,增加了植物种群内部个体之间对光合作用和养分的竞争,导致其生殖枝的生物量相对较低。而退化草地植被稀疏,无性系内植株个体间的竞争力不大,所以生殖枝也可以得到充分的养分和光合作用,所以生物量得到积累较大[34]。另外,退化草地中植物群落营养枝数量和生物量的急剧下降可能与草地土壤的风蚀、水蚀大量出现,土壤旱化、碱化、营养贫瘠加剧,造成植物群落无性系内部分株活分蘖大量死亡,造成无性系的最终死亡[35]。虽然人工恢复草地无性繁殖构件的数量和生物量均要小于未退化草地,但差距随着恢复年限的增加在逐渐减少,人工恢复草地随着生长年限的增加其无性繁殖逐渐占主导地位,表明在繁殖对策上人工恢复草地呈现出不断向天然草地靠近的态势,表明人工草地建设是恢复退化高寒草甸的一种行之有效的方法[36- 37]。
3.3 功能群水平上的植物繁殖分配对策
本研究中,从种属繁殖角度分析不同植物类群营养枝、繁殖枝数量和结构的变化可以看出,在中度、重度和极度退化草地上,莎草科的繁殖枝数占植物群落总生殖枝的比例最大,分别是48.02%,45.45%,29.07%。毒杂草次之,禾本科最少。这说明草地退化处于量变过程,但莎草科植物仍在群落中占优势地位[38]。中度和重度退化草上莎草科的繁殖枝数较高,可能是因为退化草地的高大毒杂草对嵩草属植物的一种保护,家畜不易采食[38]。对于营养枝的变化,杂类草贡献最大,造成这种现象的原因可能是一些杂类草像鹅绒委陵菜,短穗兔耳草等可以形成密集的匍匐茎网状结构,并且他们在群落中的适合度也较高[14]。3种类群的营养枝、繁殖数量和生物量都随着退化程度的增加呈现显著的降低(P<0.05)。说明草地退化明显改变植物的繁殖模式,从而影响各类群植物在整个群落中的组成比例。对于人工恢复草地而言,禾本科的营养枝和繁殖枝的生物量都显著大于莎草科和杂类草,说明人工恢复草地在一定程度上可以有效地恢复退化草地。
知识管理时代要体现以知识为核心的管理特征,高校自身具有组织、传授、整理利用知识的社会使命,所以,高校机构知识组织与管理的目标或任务,至少应该包括以下3个方面:
[1] Chen Q G. Grassland deterioration in the source region of the Yangtze-Yellow rivers and integrated control of the ecological environment. Acta Prataculturae Sinica, 2007, 1: 10- 15.
[2] Zhao X Q, Zhou H K. Eco-environmental degradation, vegetation regeneration and sustainable development in the headwaters of three rivers on Tibetan Plateau. Bulletin of the Chinese Academy of Sciences, 2005, 20(006): 471- 476.
[3] Shan Z H, Ding L L, Long R J, Ma Y S. Relationship between soil microorganisms, above-ground vegetation, and soil environment of degraded alpine meadows in the headwater areas of the Yangtze and Yellow Rivers, Qinghai-Tibetan Plateau. Acta Prataculturae Sinica, 2007, 16(001): 34- 40.
[4] Li S Y, Liu F G, Ma Y C, Niu C X. Study on status quo of grassland degradation and reasons in source region of the Three Rivers. Science and Technology of Qinghai Agriculture and Forestry, 2007,4:29- 32.
[5] Li X L. Natural factors and formative mechanism of “Black Beach” formed on grassland in Qinghai Tibetan plateau. Pratacultural Science, 2002, 19(001): 20- 22.
[6] Harris R B. Rangeland degradation on the Qinghai-Tibetan plateau: a review of the evidence of its magnitude and causes. Journal of Arid Environments, 2010, 74: 1- 12.
[7] Wu G L, Du G Z. Discussion on ecological construction and sustainable development of degraded grassland ecosystem of the Qinghai-Tibetan Plateau. Chinese Journal of Nature, 2007, 29(3):159- 164.
[8] Handel S N, Robinson G R, Parsons W F J, Mattei J H. Restoration of woody plants to capped landfills: root dynamics in an engineered soil. Restoration Ecology, 1997, 5(2): 178- 186.
[9] Provan J, Wilson S, Portig A A, Maggs C A. The importance of reproductive strategies in population genetic approaches to conversion: an example from the marine angiosperm genus Zostera. Conserv Genet, 2008, 9: 271- 280.
[10] Carlsen T M, Espeland E K, Pavlik B M. Reproductive ecology and the persistence of an endangered plant. Biodiversity and Conservation, 2002, 11: 1247- 1268.
[11] He C Q, Zhao K Y, Yu G Y. Advance in the ecological adaptability of the clonal plant in wetlands. Chinese Journal of Ecology, 1999, 18(6): 38- 46.
[12] Deng Z F, Xiao X L, Zhou X M. Primary study on reproductive strategies ofKobresiaHumilispopulation in alpine meadow. Chinese Journal of Ecology, 2001, 20(6): 68- 70.
[13] Zhang Y F, Zhang D Y. Asexual and sexual reproductive strategies in clonal plants. Frontiers of Biology in China, 2007, 2(3): 256- 262.
[14] Zhou H K, Han F, Zhou L, Zhao X Q, Liu W. Effects of alpine meadow degradation on clonal growth characteristics ofLagotisbrachystachya. Chinese Journal of Ecology, 2006, 25(8): 873- 879.
[15] Deng Z F, Zhou X M, Wang Q J. The studies of seed bank of koresia humilis meadow in Qing-Zang Plateau. Chinese Journal of Ecology, 1997, 16(5): 19- 23.
[16] Niu K C, Zhao Z G, Luo Y J, Du G Z. Fertilization effects on species reproductive allocation in an alpine meadow plant community. Journal of Plant Ecology, 2006, 30(5): 817- 826.
[17] Liu Z J, Du G Z, Chen J K. Size-dependent reproductive allocationligulariavirgaureain different habitats. Journal of Plant Ecology, 2002, 26(1): 44- 50.
[18] Li X L, Yang Y W, Zhang J, Niuzhou Y J. Growth characteristics of kobresia pygmaea clones in the “black soil beach” with different degradation. Acta Pratacultural Science, 2003, 12(003): 51- 56.
[19] Zhu Z H, Li X L, Qiao Y P, Liu W, Wang G. Study on the risk spreading strategies of clonal plant kobresia humilis under grazing selective pressures. Pratacultural Science, 2004, 21(012): 62- 68.
[20] Zhu Z H, Wang G, Zhao S L. Dynamics and regulation of clonal ramet population inkomresiahumilisunder different stocking intensities. Acta Ecologica Sinica, 1994, 14(1): 40- 45.
[21] Shang Z H, Xu P B, Ren G H, Long R J. Review of soil seed bank studies—soil seed bank function in natural ecosystem. Acta Pratacultural Science, 2006, 18(2): 175- 18.
[22] Ma Y S, Lang B N, Li Q Y, Shi J J, Dong Q M. Study on rehabilitating and rebuilding technologies for degenerated alpine meadow in the Changjiang and Yellow river source region. Pratacultural Science, 2002,19(9):1- 4.
[23] Dong M, Alateng B, Xing X R, Wang Q B. Genet features and ramet population features in the rhizomatous grass speciespsammochloavillosa. Acta Phytoecologica Sinica, 1999, 23(4): 302- 310.
[24] Zhou X M.Kobresiapygmaeameadow in China. Sicence press, 2001.
[25] Zhu L. Study on the variation of reproduction pool and soil nutrient pool of cultivated and degraded alpine grassland in the headwater areas of Qinghai-Tibetan Plateau, China. Beijing Normal University, 2011.
[26] Li J H, Pan H W, Wang G. Reproductive strategy of grassland plant. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(2): 352- 355.
[27] Liu Z M, Jiang D M, Gao H Y, Chang X L. Relationships between plant reproductive strategy and disturbance. Chinese Journal of Applied Ecology, 2003, 14(3): 418- 422.
[28] Schmid B. Clonal growth in grassland perennials: Ⅱ. Growth form and fine-scale colonizing ability. Journal of Ecology, 1985, 73: 808- 818.
[29] Li J H, Pan H W, Wang G. Reproductive strategy of grassland plant. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(2): 352- 355.
[30] Zhong Z C. Reproductive strategies of plant populations. Chinese Journal of Ecology, 1995, 14(1): 37- 42.
[31] Wang H Y, Wang Z Wen, Li L H, Chen Y J, Ren L J. Reproduction tendency of clonal plants in various habitats. Chinese Journal of Ecology, 2005, 24(6): 670- 676.
[32] Yang C, Yang L. Plasticity of clonal modules ofLeymuschinensisin response to different environments. Chinese Journal of Applied Ecology, 1998, 9(3): 265- 268.
[33] Zhou H K, Zhao X Q, Zhou L, Zhao L, Han F. Effects of different grazing intensities on growth ofpotentillaanserineclones. Acta Botanica Boreali-Occidentalia Sinica, 2006, 26(5):1021- 1029.
[34] Li X L, Zhu Z H, Yang Y W, Zhang J, Ma G X. Effects of different grazing intensities on population and clonal growth ofkobresiapygmaea. Heilongjiang Journal of Animal Science and Veterianry Medicine, 2003, 7:46- 47.
[35] Zhang J, Li X L, Yang Y W, Niuzhou Y J. The reproductive characteristics aboutkobresiapygmaeain different degraded alpine grasslands. Heilongjiang Journal of Animal Science and Veterianry Medicine, 2003, 6: 48.
[36] Wu G L, Liu Z H, Zhang L, Hu T M, Chen J M. Effects of artificial grassland establishment on soil nutrients and carbon properties in a black-soil-type degraded grassland. Plant and Soil, 2020, 333: 469- 479.
[37] Feng R Z, Long R J, Shang Z H, Ma Y S, Dong S K, Wang Y L. Establishment ofElymusnatansimproves soil quality of a heavily degraded alpine meadow in Qinghi-Tibetan Plateau, China. Plant and soil, 2010, 327: 403- 411.
[38] Xu S M. A changes of seed production of kobresia humuilis and k.pygmaea under different degenerative gradation in alpine meadow. Qinghai Prataculture, 2006, 15(1): 16- 18.
参考文献:
[1] 陈全功. 江河源区草地退化与生态环境的综合治理. 草业学报, 2007, 1: 10- 15.
[2] 赵新全,周华坤. 三江源区生态环境退化, 恢复治理及其可持续发展. 中国科学院院刊, 2005, 20(006): 471- 476.
[3] 尚占环,丁玲玲,龙瑞军,马玉寿. 江河源区退化高寒草地土壤微生物与地上植被及土壤环境的关系. 草业学报, 2007, 16(001): 34- 40.
[4] 李穗英,刘峰贵,马玉成,牛春霞. 三江源地区草地退化现状及原因探讨.青海农林科技,2007,4:29- 32. 1999, 18(6): 38- 46.
[5] 李希来. 青藏高原 “黑土滩” 形成的自然因素与生物学机制. 草业科学, 2002, 19(001): 20- 22.
[7] 武高林, 杜国祯.青藏高原退化高寒草地生态系统恢复和可持续发展探讨. 自然杂志, 2007, 29(3):159- 164.
[11] 河池全, 赵魁义, 余国营. 湿地克隆植物的繁殖对策与生态适应性. 生态学杂志, 1999, 18(6): 38- 46.
[12] 邓自发, 谢晓玲, 周兴民. 高寒草甸矮嵩草种群繁殖对策的研究. 生态学杂志, 2001, 20(6): 68- 70.
[14] 周华坤, 韩发, 周立, 赵新全, 刘伟. 高寒草甸退化对短穗兔耳草克隆生长特性的影响. 生态学杂志, 2006, 25(8): 873- 879.
[15] 邓自发, 周兴民, 王启基. 青藏高原矮嵩草草甸种子库的初步研究. 生态学杂志, 1997, 16(5): 19- 23.
[16] 牛克昌, 赵志刚, 罗燕江, 杜国祯. 施肥对高寒草甸植物群落组分种繁殖分配的影响. 植物生态学报, 2006, 30(5): 817- 826.
[17] 刘左军, 杜国祯, 陈家宽.不同生境下黄帚橐吾 (Ligulariavirgaurea) 个体大小依赖的繁殖分配. 植物生态学报, 2002, 26(1): 44- 50.
[18] 李希来, 杨元武, 张静, 牛周羊杰. 不同退化程度 “黑土滩” 高山蒿草克隆生长特性. 草业学报, 2003, 12(003): 51- 56.
[19] 朱志红, 李希来, 乔有明, 刘伟, 王刚. 克隆植物矮嵩草在放牧选择压力下的风险分散对策研究. 草业科学, 2004, 21(012): 62- 68.
[20] 朱志红, 王刚, 赵松龄. 不同放牧强度下矮嵩草 (Kobresiahumilis) 无性系分株种群的动态与调节. 生态学报, 1994, 14(1): 40- 45.
[21] 尚占环, 徐鹏彬, 任国华, 龙瑞军. 土壤种子库研究综述——植被系统中的作用及功能. 草业学报, 2006, 18(2): 175- 183.
[22] 马玉寿,郎白宁,李青云, 施建军, 董全民. 江河源区高寒草甸退化草地恢复与重建技术研究. 草业科学, 2002,19(9):1- 4.
[23] 董鸣, 阿拉腾宝, 邢雪荣, 王其兵. 根茎禾草沙鞭的克隆基株及分株种群特征. 植物生态学报, 1999, 23(4):302- 310
[24] 周兴民. 中国嵩草草甸. 科学出版社, 2001.
[25] 朱磊.三江源区高寒退化与恢复草地植被繁殖库及土壤养分库变化研究. 北京师范大学, 2011.
[26] 李金花, 潘浩文, 王刚. 草地植物种群繁殖对策研究.西北植被学报, 2004, 24(2): 352- 355.
[27] 刘志民, 蒋德明, 高红瑛, 常学礼. 植物生活史繁殖对策与干扰关系的研究. 应用生态学报, 2003, 14(3): 418- 422.
[29] 李金花, 潘浩文, 王刚. 草地植物种群繁殖对策研究.西北植被学报, 2004, 24(2): 352- 355.
[30] 钟章成. 植物种群的繁殖对策. 生态学杂志, 1995, 14(1): 37- 42.
[31] 王洪义,王正文,李凌浩,陈雅君,任丽昀. 不同生境中克隆植物的繁殖倾向. 生态学杂志, 2005, 24(6): 670- 676.
[32] 杨持, 杨理. 羊草无性系构件在不同环境下的可塑性变化. 应用生态学报,1998, 9(3): 265- 268.
[33] 周华坤, 赵新全, 周立, 赵亮,韩发. 不同放牧强度对鹅绒委陵菜克隆生长特征的影响. 西北植物学报, 2006, 26(5):1021- 1029.
[34] 李希来, 朱志红,杨元武, 张静, 马国祥.不同放牧强度下高山嵩草分株种群与克隆生长动态. 黑龙江畜牧兽医, 2003, 7::46- 47.
[35] 张静, 李希来, 杨元武, 等. 不同退化高寒草甸高山嵩草的繁殖特性. 黑龙江畜牧兽医. 2003, 6: 48.
[38] 徐世敏. 不同退化程度下高寒草甸矮嵩草与高山嵩草有性繁殖特性研究. 青海草业, 2006, 15(1): 16- 18.
Adaptationstrategiesofreproductionofplantcommunityinresponsetograsslanddegradationandartificialrestoration
LI Yuanyuan, DONG Shikui*, ZHU Lei, WEN Lu, LI Xiaoyan, WANG Xuexia
SchoolofEnvironment,BeijingNormalUniversity,Beijing100875,China
In this study, we selected the artificial grasslands with different restoration years and natural grasslands with different degradation levels to explore the changes of vegetation reproduction mode, aiming to find the feasible restoration approaches derived from the theoretical study. We found that grassland degradation and establishing artificial grassland had significant effect on the number and biomass of asexual and sexual shoots. The main conclusions are as follows: asexual reproduction dominated in the native alpine grassland. The number and biomass of asexual shoots of natural grasslands were basically lower than those of cultivated grasslands. The number of asexual shoots increased with the cultivation ages (P<0.05), and decreased with the degradation degree (P<0.05). On the contrary, the number and biomass of the sexual shoots presented opposite trend with those of the asexual shoots. With the increase of restoration years, the components of asexual and sexual shoots were close to non-degraded grassland. At the functional groups level, the number and biomass of asexual shoots were decreased with the restoration years (P<0.05), while those of sexual shoots were presented on the opposite trend; moreover, the components of grasses were significantly higher than other two groups (P<0.05). At the same time, we found that forbs accounted for absolute advantage, the similar change can be seen about degraded grassland as compared with artificial grasslands along the degradation levels. The above conclusions indicated that grassland degradation had altered the reproductive allocation and strategies about plant community, and establishing artificial grasslands is an efficient method to restore degraded alpine grasslands.
Qinghai-Tibetan Plateau; degraded grassland; artificial restoration; plant community; reproduction strategy
国家“十二五”科技支撑课题(2012BAC01B02);环保公益项目资助项目(201209033)
2012- 05- 17;
2013- 07- 20
*通讯作者Corresponding author.E-mail: dongshikui@sina.com
10.5846/stxb201205170741
李媛媛,董世魁,朱磊, 温璐, 李小艳,王学霞.青藏高原高寒草甸退化与人工恢复过程中植物群落的繁殖适应对策.生态学报,2013,33(15):4683- 4691.
Li Y Y, Dong S K, Zhu L, Wen L, Li X Y, Wang X X.Adaptation strategies of reproduction of plant community in response to grassland degradation and artificial restoration.Acta Ecologica Sinica,2013,33(15):4683- 4691.
