基于PCA的滇西北高原纳帕海湿地退化过程分析及其评价
2013-12-08杨永兴韩大勇
尚 文, 杨永兴, 韩大勇
(同济大学环境科学与工程学院污染控制与资源化国家重点实验室,长江水环境教育部重点实验室,上海 200092)
基于PCA的滇西北高原纳帕海湿地退化过程分析及其评价
尚 文, 杨永兴*, 韩大勇
(同济大学环境科学与工程学院污染控制与资源化国家重点实验室,长江水环境教育部重点实验室,上海 200092)
以滇西北高原纳帕海湖滨过度放牧、水文改变协同胁迫(简称GAD)和过度放牧单独胁迫(简称GD)退化湿地为研究对象,将其划分为4个梯度:原生湿地、轻度退化、中度退化、重度退化,采用聚类分析和主成分分析(PCA)在“植物-水体-土壤”系统上进行湿地退化过程、机制探讨及定量评价。沿退化梯度,植物群落演替规律为“水生植物群落→沼泽植物群落→沼泽化草甸植物群落→草甸植物群落”,群落结构趋于复杂,向中生植物群落演替;矿化度、总硬度、总碱度、氮、磷等水质指标在GD梯度上升,在GAD梯度下降,水文改变对水质影响显著;土壤有机质、全氮、速效氮含量降低,全磷、全钾有所增加,速效磷和速效钾含量无明显变化规律。基于PCA建立了纳帕海湖滨湿地退化模型,并给出不同程度退化湿地的阈值。
退化;过程;PCA;评价;纳帕海湿地
人类活动胁迫下湿地退化已成为当前国际湿地研究前沿领域的关键科学问题[1- 2]。过度放牧是湿地退化的主要驱动因素之一,尤其是放牧牲畜采食、践踏、排泄物污染对湿地破坏严重[3]。水源补给形式差异引起的水文改变也是导致湿地退化的关键因素[4]。深入研究过度放牧和水文改变胁迫下湿地退化过程和机制是寻求湿地生态系统开发与保护的关键。湿地退化常表现为植被、水体、土壤三大要素结构和功能的退化[5]。以往已对过度放牧胁迫下湿地生态系统植物群落演替、水质和土壤养分退化等方面有一定的深入研究[3,6- 7],对水文改变研究常侧重新构造运动下自然疏干和排水疏干、围湖造田等方面[4,8- 9]。以往研究多侧重对单一胁迫因素或部分退化指标进行湿地退化分析,对过度放牧和水文改变多因素协同胁迫下湿地退化综合研究较少[10],尤其对多因素胁迫下滇西北高原湿地“植物-水体-土壤”系统退化过程与机制研究和退化评价研究还需继续深入。为此,本文通过开展基于PCA的滇西北高原纳帕海湿地退化过程分析及其评价,旨在丰富和完善具有中国特色的高原湖滨湿地退化理论体系,为高原湿地保护与退化湿地生态恢复提供科学依据。
1 研究区域与研究方法
1.1 研究区概况
纳帕海湿地位于滇西北高原腹地,地理位置为东经99°37′—99°43′,北纬27°49′—27°55′,海拔3260m,行政上隶属于云南省迪庆藏族自治州。该区年均温度5.4℃,最冷月平均温度为-3.8℃,最热月平均温度为13.2℃,极端低温-25.4℃,极端高温24.5℃。该区属金沙江水系,年均降水量619.9 mm,受西南季风影响每年6、7月份形成大量降水,湖面面积达到3125hm2。但8月后湖水退落,10月前后因秋季季风退缩再次产生降雨,湖水再次上涨并于11月后退落。湖水退落后湖面大幅度缩小,湖滨带广阔,发育了大面积沼泽湿地和沼泽化草甸。该区植物物种十分丰富,经初步统计共有植物115种,隶属38科、82属,主要包括水葱群落(Com.Scirpusvalidus)、水蓼群落(Com.Polygonumhydropiper)、刘氏荸荠群落(Com.Eleocharisliouana)、鹅绒委陵菜群落(Com.Potentillaanserine)、发草-矮地榆群落(Com.Deschampisacaespitosa-Sanguisorbafiliformis)等15个群落类型[11]。该区主要土壤类型为沼泽土、泥炭土、沼泽化草甸土[12]。纳帕海湿地牦牛、黄牛、马、猪等牲畜过度放牧现象严重,实际载畜量约为6000羊单位,超过理论载畜量132.5%[13],过度放牧引起湿地大面积退化,甚至出现沙化现象[12]。纳帕海修建的环湖公路和周边村民砌筑的道路改变了湖滨湿地水文地貌,导致湿地景观破碎化和生态功能退化[14]。
1.2 研究方法
1.2.1 样地选择
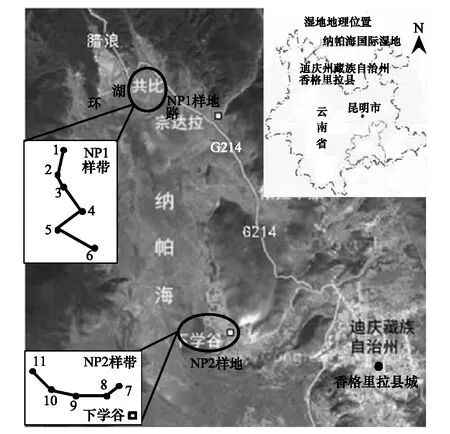
图1 研究区地理位置及样地设置Fig.1 Geographic position of study area and plot establishment
根据典型性、代表性和可行性原则,选取纳帕海湖滨湿地2个退化区域建立两条样带(编号分别为NP1和NP2样带),设置退化样点(图1)。NP1样带位于纳帕海湖滨宽广沟谷湿地,因修建道路导致湿地和湖泊隔断,水源补给主要来自地下水、融雪水和降水,牲畜放牧严重;NP2样带位于宽广湖滨的谷地,谷地下部湿地受周期性水位上涨时湖水补给,谷底上部区域受融雪水和降水补给,是牲畜主要放牧区。NP1样带作为受水文改变和过度放牧协同胁迫退化样带(简称GAD),自沟谷陡坡至湖滨带设置6个不同程度退化样点,编号为1—6;NP2样带作为主要受过度放牧胁迫退化样带(简称GD),自湿地边缘至湖滨带设置5个不同程度退化样点,编号为7—11。
1.2.2 样品采集和分析
于2010 年7—8 月在植物生长季进行野外生态观测与样品采集。植物群落生态观测采用样方法,每个样点设置2 个1m×1m 植物群落样方,统计植物物种,群落植物盖度、高度和密度。采用刈割法收获样方内植物地上生物量称其鲜质量,杀青并烘干处理后称其干质量。使用自制采样器采集上层水样,地表无水处采取经地表径流和降水汇入人工挖坑后的积水,测定矿化度、总硬度、总碱度、总氮、氨氮、硝态氮、总磷等[15]。土壤样品采集使用探坑法,根据土壤发生层(草根层、腐殖质层或泥炭层、潜育层和母质层)顺序从上到下采样深度控制在0—10cm、10—30cm、30—50cm和50—80cm,部分层次适当调整,每个土壤发生层采集土样2份,每份质量在1kg左右。对土壤样品进行pH、有机质、全氮、全磷、全钾、速效氮、速效磷、速效钾测定[16]。样品分析化验使用国家标准方法,采用平行样测定,指标分析精度均在标准允许误差之内。
1.2.3 数据处理
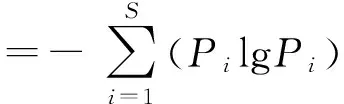
通过聚类分析对样点进行退化分类和分级;基于主成分分析(PCA)揭示湿地退化特征因子,选取前两个主成分为坐标轴做指标排序图,并建立基于简化后主成分的多元回归模型,计算湿地退化梯度综合指标分值(PC值),划分出分值区间,每个分值区间代表了相应的退化梯度,以此定量描述湿地退化情况,并进行湿地退化评价。
使用Microsoft office 2007软件进行数据分析和作图;使用BioDiversity pro软件计算物种多样性;使用SPSS 13.0软件完成聚类分析和主成分分析。
2 结果与分析
2.1 湿地退化分类与分级
遵循生态学原则,在GAD和GD退化梯度上共11个样点分别选取群落类型指数、微地貌指数、湿地积水深度、距湖岸带距离、干扰强度、植物残体层厚度、群落盖度、群落植物平均高度、样方内物种数目、水生植物重要值、沼生植物重要值、湿生植物重要值、中生植物重要值、土壤pH、土壤有机质含量、水体总氮、总磷含量共17个典型指标作为湿地退化分类要素。并结合自然地理特征、生态环境特征和湿地发育程度,将其中的定性指标作定量化处理并赋值(表1)。其中,微地貌指数是按地貌对湿地发育影响程度赋值;干扰强度赋值区间从0 到10,依次代表无干扰的原生湿地到极度退化出现裸地的湿地退化程度。
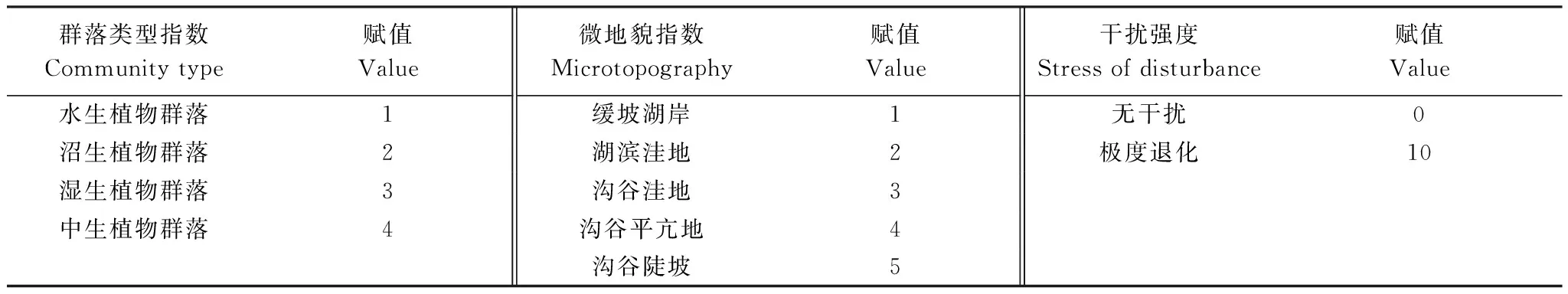
表1 纳帕海湖滨湿地退化指标定量化处理
微地貌指数是按地貌对湿地发育影响程度赋值;干扰强度赋值区间从0到10,依次代表无干扰的原生湿地到极度退化出现裸地的湿地退化程度
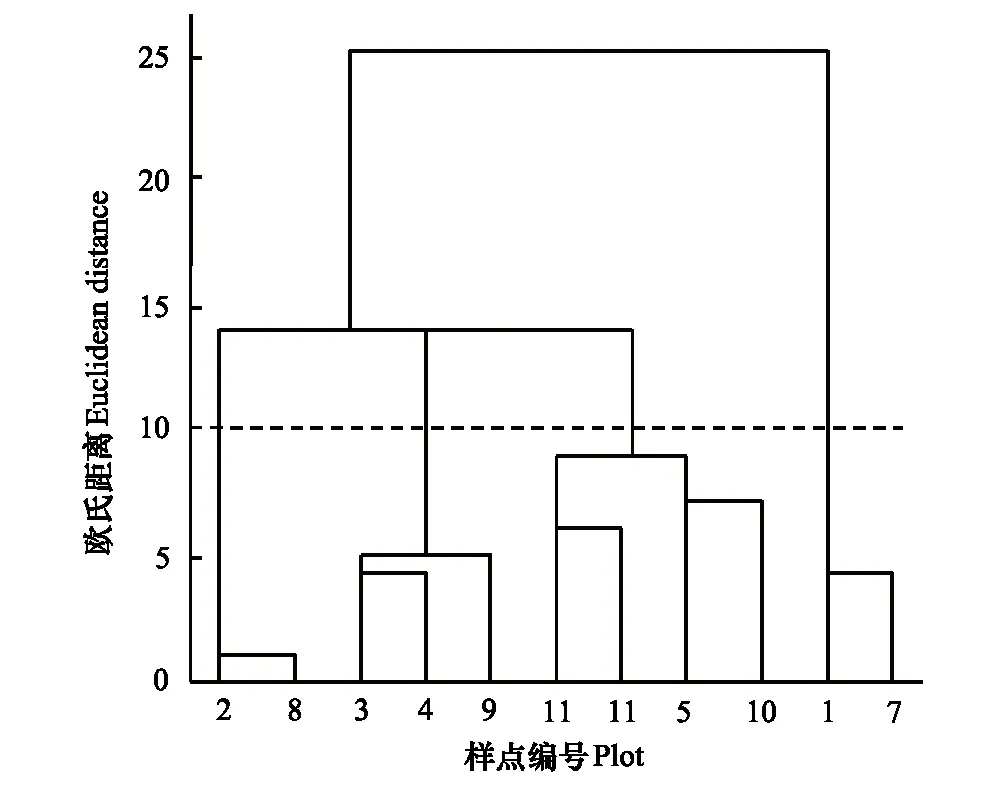
图2 纳帕海湖滨湿地退化样点聚类分析 Fig.2 Hierarchical cluster analysis of degraded samples in lakeside wetlands, Napahai
聚类分析结果如图2。一方面为了保证尽量细分退化样点以利于深入揭示退化过程,另一方面要保证分类后各组之间特性差异明显,故在图中选取欧氏距离为10,可以将11个退化样点分为4组:第1组包含样点2、8;第2组包含样点3、4、9;第3组包含样点5、6、10、11;第4组包含样点1、7。根据聚类分析结果,并结合样点环境特征,将2个样带共计11个样点分为4个退化梯度:原生湿地(PW)、轻度退化(LW)、中度退化(MW)、重度退化(SW) (表2)。
2.2 湿地植被退化特征分析
样方统计显示, GAD退化梯度上植物群落共包含高等植物24种,隶属14科22属。其中莎草科(Cyperaceae)4属4种、毛茛科(Ranunculaceae)3属3种、蓼科(Polygonaceae)1属3种、伞形科(Umbelliferae)2属2种、禾本科(Poaceae)2属2种、蔷薇科(Rosaceae)2属2种,其它为十字花科(Brassicaceae)、车前科(Plantaginaceae)、灯心草科(Juncaceae)、睡菜科(Menyanthaceae)、玄参科(Scrophulariaceae)、唇形科(Lamiaceae)、龙胆科(Gentianaceae)、眼子菜科(Potamogetonaceae)各1属1种。GD退化梯度上植物群落共包含高等植物26 种,隶属15科25属。其中莎草科4属4种、毛茛科5属5种、菊科(Compositae)2属3种、玄参科2属2种、蝶形花科(Papilionaceae)2属2种,其它为十字花科、车前科、大戟科(Euphorbiaceae)、灯心草科、禾本科、黑三棱科(Sparganiaceae)、蔷薇科、伞形科、杉叶藻科(Hippuridaceae)、睡菜科各1属1种。
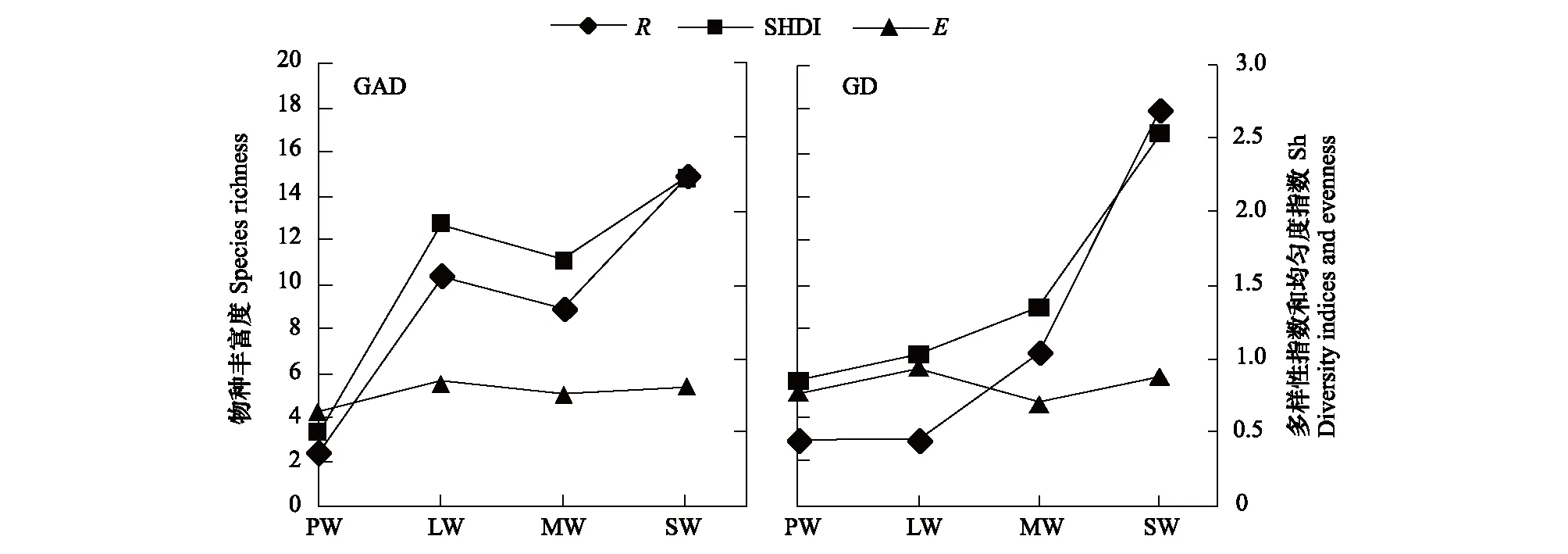
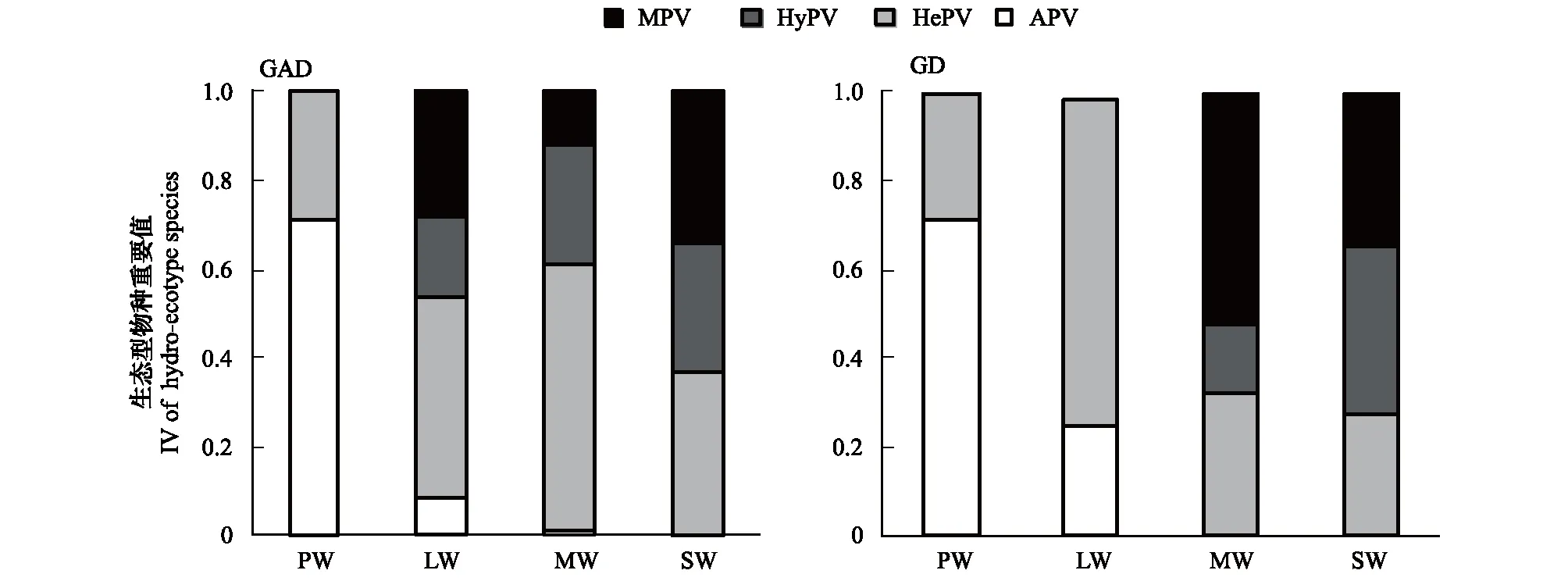
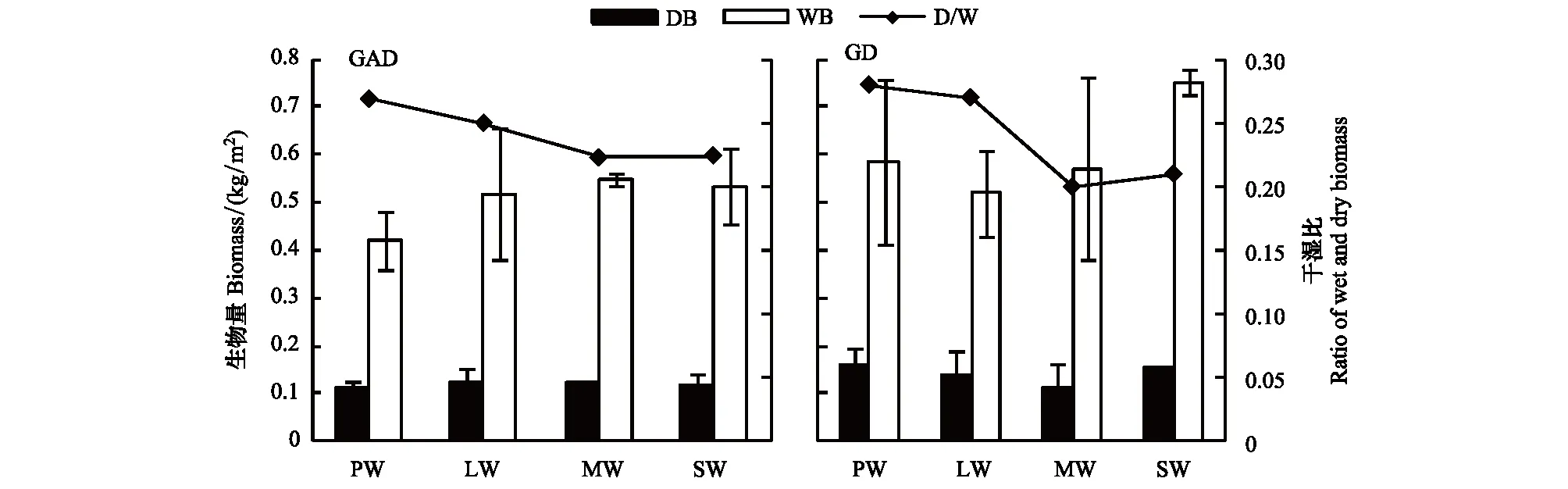
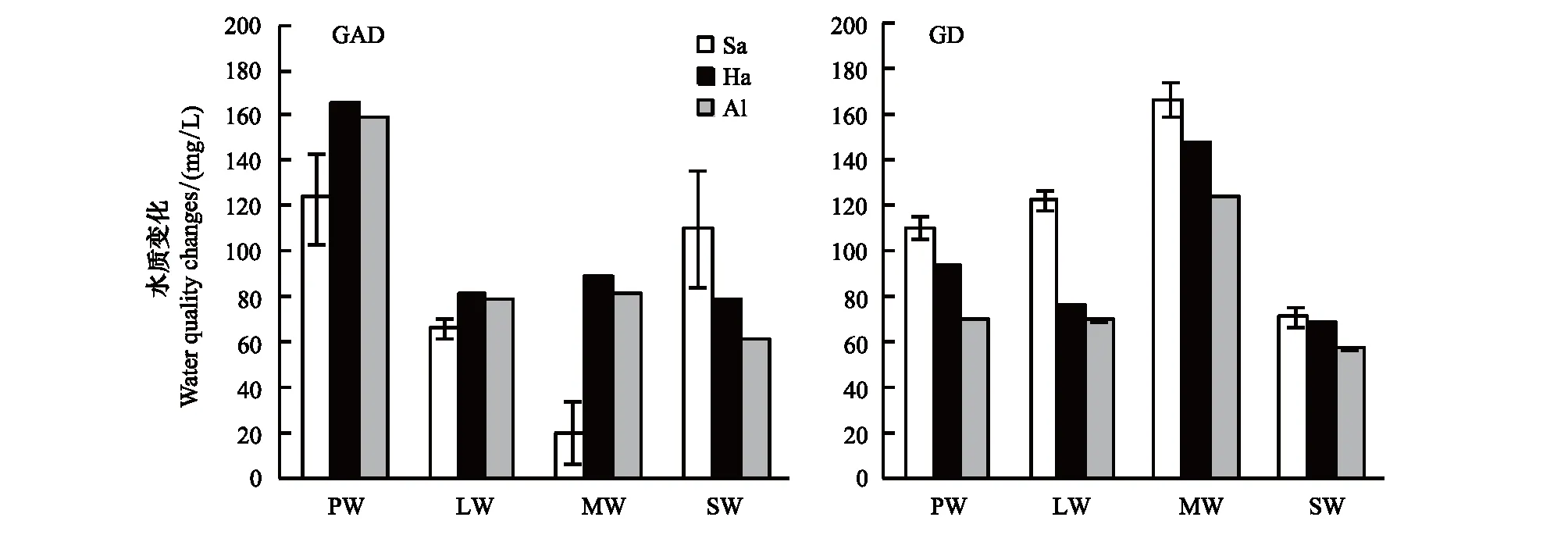
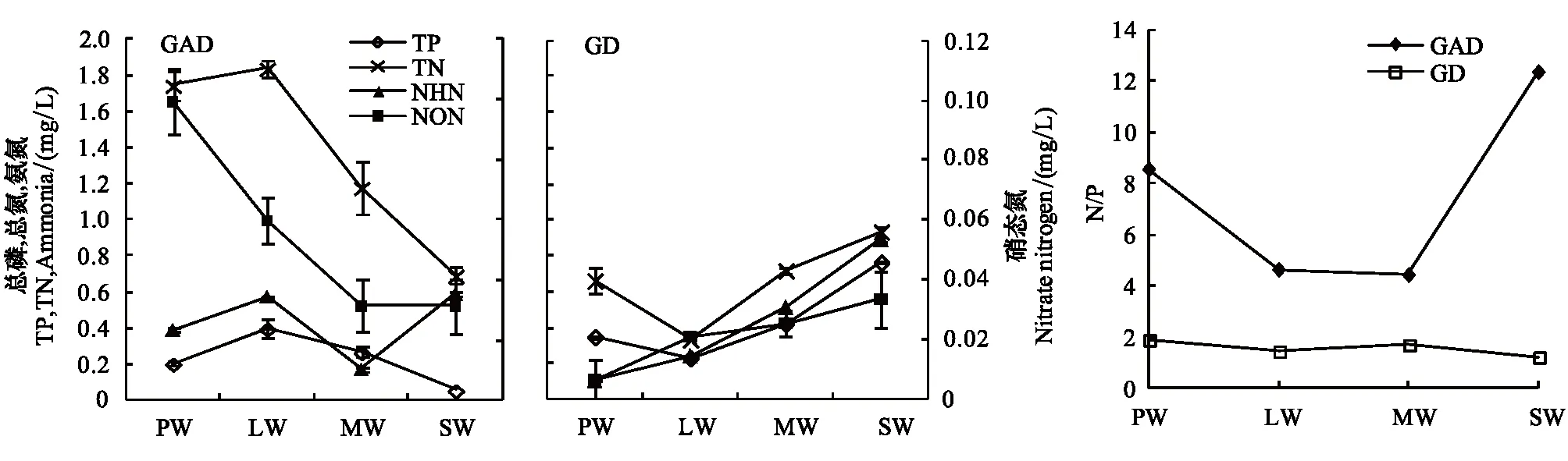
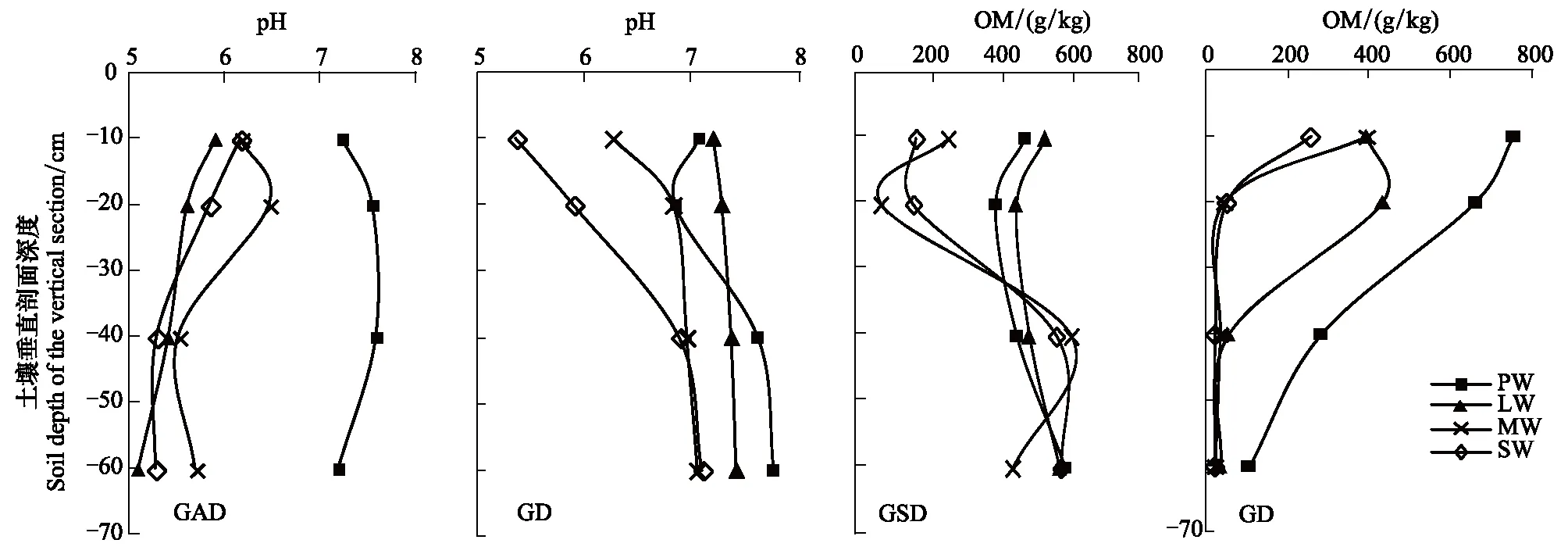
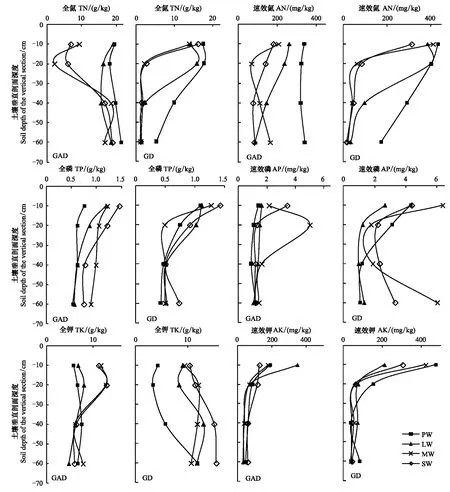
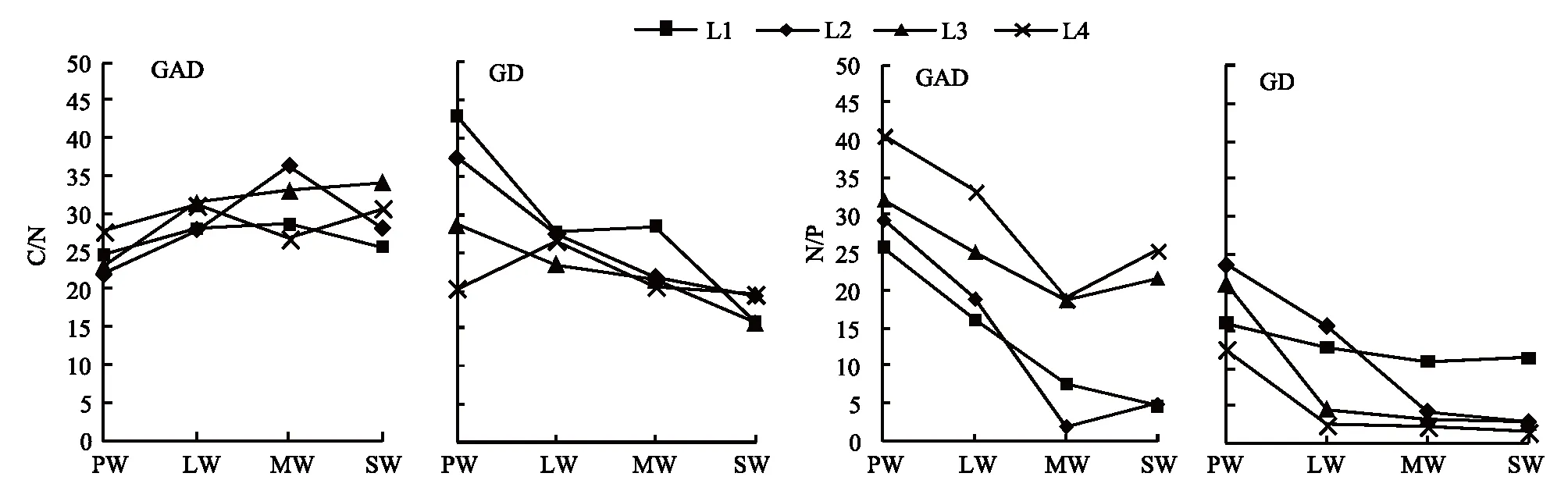
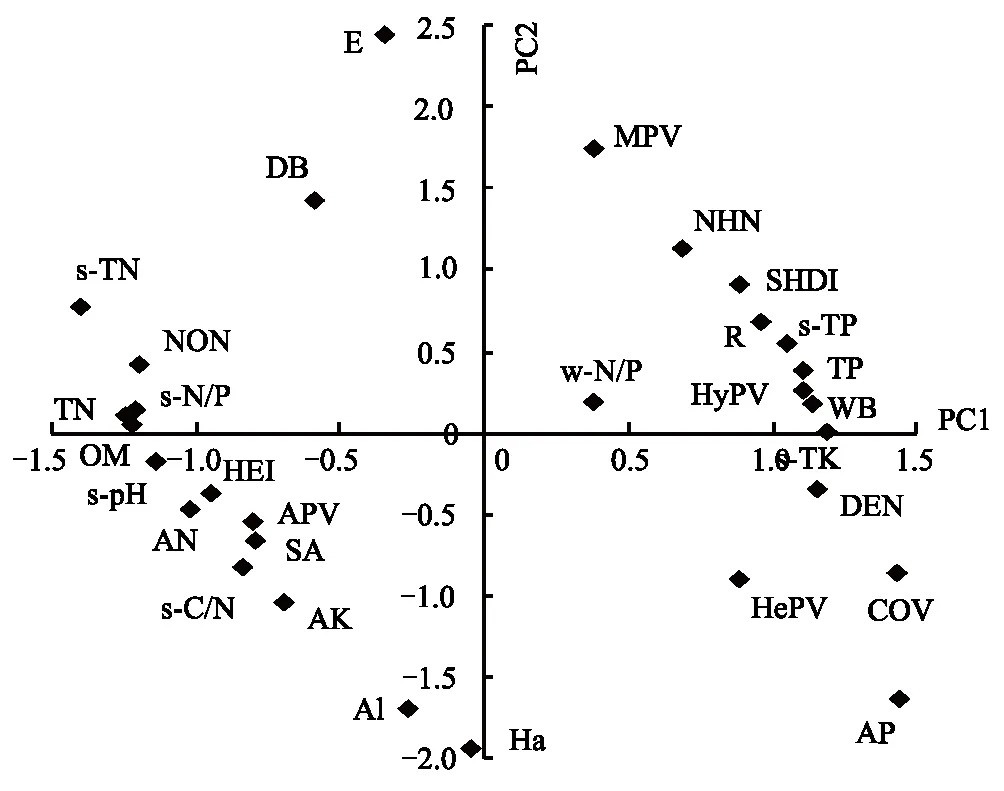
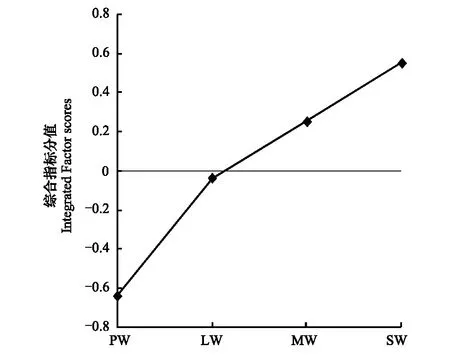
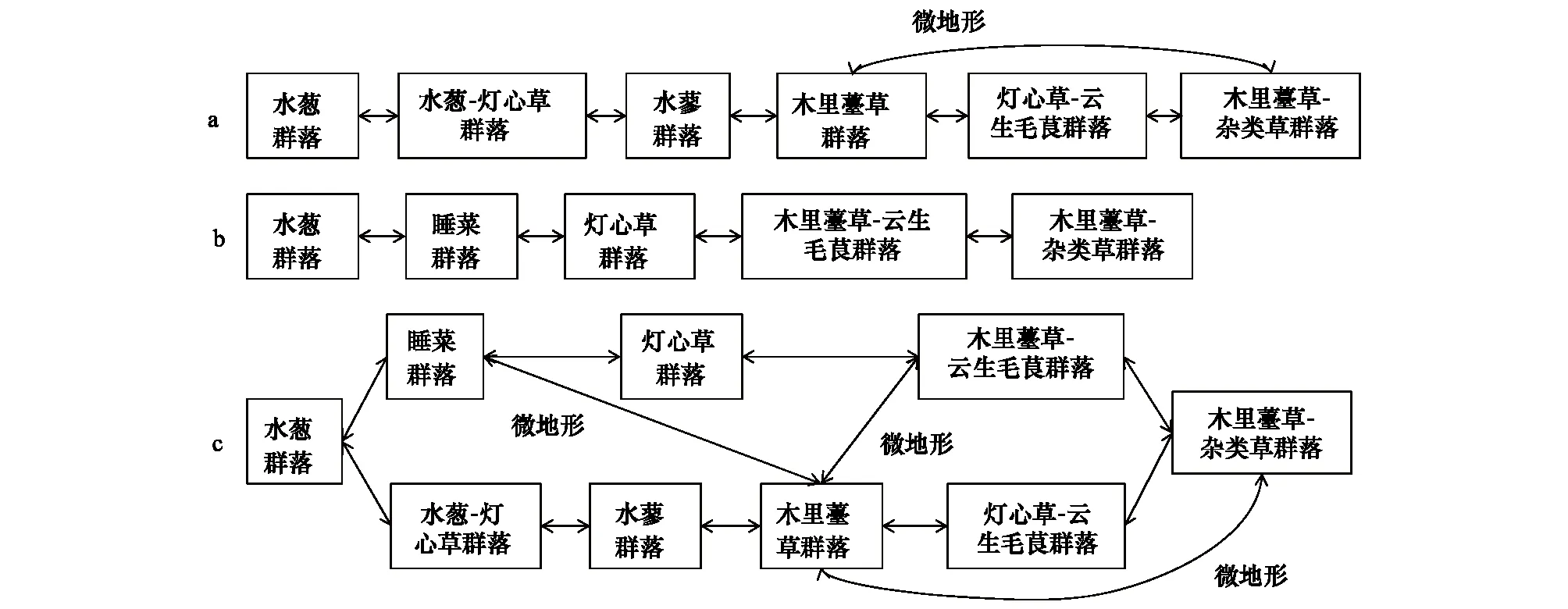
GAD和GD退化梯度上植物科、属、种数目逐渐增加,群落高度下降,向低矮型中生群落演替,群落密度和群落盖度也逐渐增加。随退化加剧,多样性指数和物种丰富度均表现出逐渐增加趋势。但GD退化梯度上LW出现草丘,伴生较多中生植物,导致LW的物种多样性稍有增加,多样性指数和物种丰富度排序为PW 退化梯度上,水生植物逐渐退出群落,沼生植物和湿生植物开始增加,最后以中生植物居多,成为群落主体。SW下中生植物种数占物种总数目的比例也较高,GAD和GD这一比例分别达46.7%和61.1%(图4)。 表2 纳帕海湖滨湿地退化样点环境特征 PW: 原生湿地, Primary wetland; LW: 轻度退化, Lightly degraded wetland; MW: 中度退化, Moderately degraded wetland; SW: 重度退化, Severely degraded wetland 图3 纳帕海湖滨湿地植物群落物种多样性变化Fig.3 Plant species diversity along the degraded gradients of lakeside wetlands, NapahaiR: 物种丰富度Species richness; SHDI: 多样性指数Shannon-Wiener diversity indices; E: 均匀度指数Evenness; GAD: 过度放牧、水文改变协同胁迫, the synergistic impacts of overgrazing stress and water regime; GD: 过度放牧单独胁迫, the single impacts of overgrazing stress; PW: 原生湿地, Primary wetland; LW: 轻度退化, Lightly degraded wetland; MW: 中度退化, Moderately degraded wetland; SW: 重度退化, Severely degraded wetland 图4 纳帕海湖滨湿地退化梯度上植物水分生态型特征 Fig.4 Plant hydro-ecotypes along the degraded gradients of lakeside wetlands, NapahaiAPV: 水生植物重要值IV of aquatic plant; HePV: 沼生植物重要值IV of helophyte plant; HyPV: 湿生植物重要值IV of hygrophyte plant; MPV: 中生植物重要值IV of mesophyte plant 水生植物群落向湿生、中生植物群落演替过程中,湿地生产力逐渐提高,沿退化梯度,群落地上生物量(鲜质量)逐渐增加,地上生物量(干质量)变化趋势不明显。GAD退化梯度上生物量(鲜质量)增幅达27.3%,GD上生物量(鲜质量)先稍有下降,然后逐渐上升,整个GD上增幅达28.7%。而地上生物量(干质量)在两个退化梯度上增幅不大。生物量干湿比在两个退化梯度上均呈下降趋势,降幅分别达16.6%和25.0%(图5)。 图5 纳帕海湖滨湿地退化梯度上群落生物量变化 Fig.5 Changes of plant biomass along the degraded gradients of lakeside wetlands, NapahaiDB: 地上生物量(干质量)Dry biomass; WB: 地上生物量(鲜质量)Wet biomass; D/W: 干湿比Ratio of wet and dry biomass 2.3 湿地水体退化特征分析 2.3.1 水体矿化度、总硬度、总碱度变化特征 GAD退化梯度受水文改变影响,导致湿地以高矿化度、硬度和碱度的地下水补给为主,而MW和SW的地表无积水,补给形式主要为融雪水和降水,故GAD退化梯度上PW的水体矿化度、总碱度和总硬度偏高,然后随退化加剧逐渐降低。而GD梯度上,随着过度放牧加剧,牲畜粪便排泄物输入导致水质恶化,牲畜践踏、啃食、翻拱等都不同程度地造成地表裸露,土壤矿化作用增强,导致受纳水体矿化度、总硬度和总碱度升高,SW的各指标值偏低(图6)。 图6 纳帕海湖滨湿地退化梯度上水体基本性质变化 Fig.6 Changes of water properties along the degraded gradients of lakeside wetlands, NapahaiSa: 矿化度Salinity; Ha: 总硬度Total hardness; Al: 总碱度Total alkalinity 2.3.2 水体氮、磷指标变化特征 GAD退化梯度上,水体总氮、硝态氮和总磷含量呈下降趋势,氨氮含量有所波动。水体总氮降幅达60%,硝态氮降幅达到68%。总磷呈先上升后下降的变化趋势。GD上总氮、氨氮、硝态氮、总磷均呈上升趋势。其中硝态氮上升了5.1倍,氨氮上升了8.6倍。总氮含量先下降然后逐渐上升,SW水体总氮含量是PW的1.4倍,总磷含量增长超过2倍。GD退化梯度上氨氮含量占总氮的比例很高,MW和SW时这一比例分别达到71%和95%。SW的水体总氮含量(0.933mg/L)接近国家地表水环境质量标准(GB3838—2002)中Ⅲ类水总氮标准,总磷含量(0.762mg/L)远远超过Ⅴ类水总磷标准(图7)。 随退化加剧,GAD的水体N/P呈先下降后上升趋势,从PW的8.5∶1下降到4.6∶1,然后在SW又增加到12.3∶1。GD下N/P变化幅度不大,稳定在1.4∶1—1.9∶1,但低于GAD(图7)。对比可知,GAD的水体总氮含量均高于GD,导致GAD退化梯度上水体N/P远远高于GD,二者受水源补给方式的差异导致水质变化较明显。 图7 纳帕海湖滨湿地退化梯度上水体氮、磷含量变化 Fig.7 Changes of water N, P content along the degraded gradients of lakeside wetlands, NapahaiTN: 总氮Total nitrogen; NHN: 氨氮Ammonia nitrogen; NON: 硝态氮Nitrate nitrogen; TP: 总磷Total phosphorus 2.4 湿地土壤退化特征分析 2.4.1 土壤pH和有机质变化特征 纳帕海湿地土壤以中性偏酸性土壤为主,GAD上原生湿地土壤pH要高于退化后湿地,但LW的土壤pH在整个退化梯度上最低,表层和亚表层土壤pH排序为:LW 沿退化梯度,受土壤矿化作用增强和人为干扰加剧影响,土壤有机质(OM)含量呈下降趋势。GAD退化梯度上表层土壤OM呈下降趋势,下层OM无明显变化规律。沿GD退化梯度,PW的土壤OM含量较高,表层和亚表层均高于下层。随退化加剧,GD的土壤OM含量呈下降趋势(图8)。 图8 纳帕海湖滨湿地退化梯度上土壤pH、有机质变化 Fig.8 Changes of soil pH and OM along the degraded gradients of lakeside wetlands, NapahaipH: 土壤pH Soil pH; OM: 土壤有机质Organic matter 2.4.2 土壤养分指标变化特征 GAD退化梯度上,土壤全氮含量逐渐下降,特别是表层(0—10cm)和亚表层(10—30cm)下降趋势较为明显,降幅分别高达65.1%、67.1%。全磷和全钾却随退化加剧呈逐渐上升趋势,表层和亚表层全磷增幅分别达91.8%和95.9%,全钾在PW土壤表层和亚表层含量均增加1倍左右。速效氮在GAD上逐渐下降,表层降幅达到46.4%,亚表层降幅达56.6%。速效磷含量随退化加剧有所升高,表层增幅明显。速效钾含量变化趋势并不明显,表层速效钾含量排序为SW GAD土壤垂直剖面上,全氮含量随深度增加有所上升,PW和LW的全氮含量随垂直深度增加而有少量增加,但MW和SW时增加较明显。全磷、全钾含量随深度增加有所下降,在SW时下降最为明显,50—80cm处全磷、全钾含量分别只有表层的51.3%和53.4%。速效氮和速效磷含量随土壤深度增加下降趋势不大,但速效钾含量下降幅度较大,在30cm以下土壤速效钾含量只为表层的11%—46%,具有明显的表聚性特征。 GD退化梯度上,土壤全氮含量呈下降趋势,PW向MW退化时全氮含量下降较明显,表层全氮下降幅度为21.1%,亚表层达到88.1%。全磷和全钾含量却随退化加剧而逐渐升高,表层土壤全磷含量增幅达27.9%,表层全钾含量增幅高达143.1%。速效氮含量呈下降趋势,表层降幅明显。土壤速效磷在整个GD上上升幅度并不明显,表层土壤速效磷含量排序为LW GD土壤垂直剖面上,全氮主要集中在表层(0—10cm)和亚表层(10—30cm),全磷含量在30cm深度以下基本上保持在0.5g/kg水平,显著低于上层。但是全钾含量却随土壤深度增加有所升高。土壤速效养分具有明显的表聚性,特别是速效钾。垂直剖面上MW和SW的速效氮下降趋势比LW和PW明显,MW土壤30cm以下速效氮含量是表层的10%左右,而SW时也只有15%。速效磷含量在50—80cm会高于中间10—50cm层,但仍低于表层0—10cm处。速效钾在30cm以下含量已降低到表层的1/5—1/8(图9)。 图9 纳帕海湖滨湿地退化梯度上土壤氮磷钾养分沿垂直剖面变化Fig.9 Changes of N, P, K content in the soil vertical profiles along the degraded gradients of lakeside wetlands, Napahai GAD上土壤C/N在各层土壤中呈上升趋势,而在GD上C/N呈逐级下降趋势。GD退化梯度PW及LW的C/N较高,退化加剧后则逐渐降低,各土壤层均表现出下降趋势。GAD和GD上N/P逐渐下降,SW时略有升高,总体呈下降趋势。GAD的下层N/P要高于上层,特别是第四层N/P明显高于上层。而在GD上正相反,上层N/P基本要高于下层(图10)。在两个退化梯度上土壤C/N和N/P主要取决于全氮含量的变化,氮素的赋存形态和含量是湿地退化的最明显特征。 图10 纳帕海湖滨湿地退化梯度上土壤碳氮比、氮磷比变化 Fig.10 Changes of soil C/N and N/P along the degraded gradients of lakeside wetlands, NapahaiL1: 表层 (0—10cm); L2: 亚表层 (10—30cm); L3: 第三层(30—50cm); L4: 第四层 (50—80cm) 图11 纳帕海湖滨湿地系统指标PCA排序图 Fig.11 PCA ordination diagram of indices in degraded wetlands on the lakeside zone, NapahaiCOV:群落盖度 Coverage; HEI: 群落高度Height; DEN: 群落密度Density. 其他指标同前The others are same to before 对30个“植物-水体-土壤”指标采用Z标准化(标准值=[原始值-原始值均值]/原始值标准差)处理后进行主成分分析,前3个主成分分别解释了54.8%、29.1%和15.9%的原始信息,累计解释量为99.8%。其中第一、第二主成分共携带83.9%的原始信息,选择前两个主成分为坐标轴做系统指标排序散点图(图11)。第一主成分(PC1)与土壤速效磷、植物群落盖度、土壤全钾相关性最高,与土壤全氮、水体总氮、土壤有机质、土壤氮磷比相关性最低;第二主成分(PC2)与植物群落均匀度指数、中生植物重要值、地上生物量(干质量)的相关性最高,与水体总硬度、总碱度、土壤速效磷、速效钾、水生植物重要值、群落盖度相关性最低。因此,第一主成分集中代表了土壤信息,主要反映土壤指标变化。第二主成分集中代表了植物群落信息,主要反映植物群落指标变化。 通过主成分分析,将30个“植物-水体-土壤”指标简化为三个主成分,分别携带了原始指标54.8%、29.1和15.9%的信息,基于PCA三个主成分建立多元回归模型: PC=0.548×PC1 + 0.291×PC2+0.159×PC3 计算湿地退化梯度综合指标分值(PC值),定量描述滇西北高原湖滨纳帕海湿地退化程度。结果显示(图12),沿退化梯度,PC值呈逐级上升趋势,PW的PC值为负值,然后在MW和SW时为正,PC值越大,代表湿地退化越严重。结合野外观察和PCA分析,将PC值划分为4个区间建立湿地评价模型:(1)PC≤-0.300,为原生湿地(PW);(2)-0.300 图12 纳帕海湖滨退化湿地综合指标分值 Fig.12 Integrated factor scores of five degraded gradients of lakeside wetlands, Napahai 4.1 湿地植被退化过程及机制 随湿地退化加剧,植物群落物种科、属、种数目逐渐增多,群落密度和盖度增加,群落高度降低,地上生物量(鲜质量)有所增加。物种丰富度和多样性指数逐渐增加。水生植物逐渐退出生境,沼生、湿生植物比例逐渐增加,最后中生植物开始占据群落的主体。纳帕海湿地过度放牧胁迫下植被退化较快,适口性植物比例严重下降,鹅绒委陵菜(Potentillaanserina)、北水苦荬(Veronicaanagallis)、矮地榆(Sanguisorbafiliformis)等杂草类比例较高,并出现草甸典型退化植物大狼毒(Euphorbiajolkinii)等,部分区域甚至出现裸地,过度放牧在一定程度上加速了向草甸演化这一过程。前人对若尔盖高原沼泽植被退化研究时也发现这一规律,过度放牧干扰了群落固有演替模式,在演替方向上未发生本质变化,但过度放牧起到加速沼泽植被演替的作用[18]。 经过两年湖滨带湿地野外考察与植物样方观测,并结合地貌、水文、土壤及环境条件,得到滇西北高原纳帕海湖滨湿地植物群落演替模式(图13)。GAD样带受水文改变胁迫,水分梯度影响群落演替模式,从积水较深的水葱群落向水葱-灯心草群落、水蓼群落演替,然后随地表积水减少向木里薹草群落退化,严重退化的草甸区群落退化成灯心草-云生毛茛群落和木里薹草-杂类草群落(图13a)。GD样带所处区域地势较缓,具有渐变生境特征,湖滨带主要以水葱群落为主,然后逐渐向睡菜群落演替,沼泽化草甸主要以灯心草群落为主,然后逐渐向木里薹草-云生毛茛群落、木里薹草-杂类草群落演替,严重退化区地表甚至无植被(图13b)。在纳帕海湿地综合演替模式上,水分梯度主导该区植物群落演替,微地貌变化和人为活动干扰加剧会导致湿地植物群落出现跳跃式演替,但并不改变群落演替方向(图13c)。在前人的研究基础上进一步深入后[13,19],总结出纳帕海湿地植物群落演替规律为:水生植物群落→沼泽植物群落→沼泽化草甸植物群落→草甸植物群落。 图13 纳帕海湖滨湿地植物群落演替模式 Fig.13 Plant community succession of the lakeside wetlands community, Napahaia: GAD退化梯度样带Transect of GAD; b: GD退化样带Transect of GD; c: 纳帕海湿地综合演替模式General succession mode of wetlands 4.2 湿地水体退化过程及机制 在GAD和GD上,水体指标变化规律有所差异。沿GAD退化梯度,原生湿地水体矿化度、总硬度、总碱度较高,随退化加剧逐渐下降。总氮、硝态氮和总磷指标在整个GAD退化梯度呈下降趋势,氨氮指标出现波动。沿GD退化梯度,矿化度、总硬度、总碱度、总氮、氨氮、硝态氮、总磷等指标均逐渐升高。且氨氮含量占总氮的比例也逐渐升高,水体呈现一定富营养化趋势。 GAD与GD上水体指标变化趋势有所差异,这与湿地退化胁迫因素有紧密联系。GD退化主要受过度放牧影响,沿退化梯度水质氮、磷指标有所升高,主要原因是:(1)过度放牧胁迫下牛、马、猪等牲畜排泄物过量输入湿地水体,有机物容易分解;(2)牲畜践踏导致土壤结构功能破坏,有机物分解加快使土壤氮、磷营养物进入沼泽地表水体,总氮和总磷严重超标;(3)湿地退化后,净化水质能力下降,环境容量减小,水质会随之退化,过度放牧胁迫下湿地水体退化是一个正反馈的过程。而GAD退化受水文改变影响导致湿地水源补给主要来自高矿化度、高硬度和高碱度的地下水,故原生沼泽湿地水体指标含量较高;而重度退化时湿地地表无积水,主要受融雪水和降水补给,沿退化梯度水体指标有所降低。湿地水体的营养程度会对植物群落演替产生重要影响[20],而水文改变胁迫对水质退化影响较明显。 4.3 湿地土壤退化过程及机制 沿退化梯度,土壤pH、有机质、全氮含量逐渐下降,其机制如下:湿地逐渐向草甸退化,土壤通气性增加,加速了有机质矿化过程。植物群落啃食严重导致有机物输入量减少,分解量增加,土壤有机质含量急剧下降。土壤全氮与土壤有机质之间存在显著正相关性,放牧胁迫下土壤全氮亦随分解加剧而降低。研究发现,随着湿地土壤水分被排干,通气性增加导致全氮被分解而使速效氮含量增加[12],但本研究中土壤速效氮含量却随沼泽土向沼泽化草甸土退化过程中逐渐下降。分析可知,采样期7、8月为该区植物生长旺季,速效氮被植物吸收和微生物利用增强而有所下降。前人对纳帕海湿地土壤速效氮含量研究也发现,沼泽土速效氮组分含量显著高于沼泽化草甸土和中生草甸土[21]。 湿地土壤钾含量主要来自可利用营养物的累积,它取决于湿地植物吸收和植物残体死亡后累积[22]。退化梯度上群落生产力逐渐提高,土壤全磷、全钾含量表现出逐渐上升趋势。而土壤速效磷养分上升趋势并不明显,速效钾养分变化无明显规律。此外,GD退化梯度土壤碳氮比和氮磷比沿湿地退化梯度逐渐下降,GAD退化梯度上土壤氮磷比也逐渐下降,但碳氮比稍有升高,这主要受土壤全氮含量影响。有对美国佛罗里达州大沼泽地湿地退化研究发现,其退化原因是沼泽区水文条件和土地利用方式变化以及过量施肥导致沼泽磷污染严重,进而引起湿地退化[23]。随退化加剧,土壤有机质和氮磷含量受影响较大[24],尤其是对氮含量的影响最显著,氮素的赋存形态和含量变化是湿地退化的最明显特征。 在土壤垂直剖面上,GD退化梯度土壤全氮、速效氮养分比GAD表聚性更明显。GD由于受湖水周期性补给,氮素在干湿交替下分解较为严重,植物残体层厚度要低于GAD退化梯度,下层土壤全氮和速效氮含量较低。而严重退化后土壤养分积累量急剧下降,植物残体厚度非常薄,下层养分含量明显低于表层,湿地退化后土壤逐渐变得贫瘠,这与三江平原典型湿地土壤营养元素空间分异规律相近[25]。 4.4 湿地指标排序分析 对多指标进行PCA分析,第一主成分主要代表土壤指标,第二主成分主要代表植物指标。随湿地退化加剧,退化梯度与土壤指标的相关性逐渐增加,湿地退化对土壤指标影响显著。湿地土壤养分变化反映了土壤环境的改变,并随胁迫强度的加剧而退化。在人类活动干扰下,土壤养分含量降低,湿地土壤退化日趋严重,人类活动是导致土壤质量下降的主要原因,与我国三江平原、若尔盖高原以及国外湿地土壤退化形成机制基本相同[25- 27]。原生湿地退化梯度与植物指标呈负相关,中度、重度退化梯度与植物指标相关性逐渐增加,说明随湿地退化加剧植物群落演替速率逐渐加快,由水生或沼生植物群落向中生植物群落退化。 相对于土壤退化和植被退化对纳帕海湖滨湿地退化的响应程度,水体指标受湿地退化影响复杂。一方面是由于水文周期变化导致湿地水质并不十分稳定;另一方面,严重退化时湿地已经完全退化为草甸,地表已无明显积水,水源补给方式差异对水质影响程度较大。 (1)随湿地退化加剧,植物群落结构趋于复杂,物种多样性、丰富度增加,地上生物量也逐渐增加。水生、沼生植物群落逐渐向湿生、中生植物群落演替。纳帕海湖滨湿地植物群落演替模式为:水生植物群落→沼泽植物群落→沼泽化草甸植物群落→草甸植物群落。水文改变影响下微地形差异导致湿地植物群落类型更加复杂。放牧活动干扰了群落固有演替过程,在演替方向上未发生本质变化,但加快群落演替速率,在一定程度上加速了沼泽化草甸向草甸演替这一过程。 (2)过度放牧胁迫下水体矿化度、总硬度、总碱度升高,氮、磷含量增加,而水文改变和过度放牧协同胁迫对水体影响规律与之相反,水文改变是导致湿地水质退化规律差异的主要因素。 (3)过度放牧和水文改变胁迫导致土壤养分含量逐渐下降,土壤有机质、全氮、速效氮含量降低,土壤全磷、全钾增加,速效磷、速效钾含量变化不明显。在垂直剖面上,土壤全磷、速效磷和速效钾含量具有表聚性。过度放牧干扰对土壤结构和养分变化负面影响更剧烈。氮素的赋存形态和含量是湿地退化的最明显特征。 (4)PCA研究显示,植物群落和土壤指标随着纳帕海湖滨湿地退化加剧变化显著,而水质的响应程度复杂。基于PCA建立纳帕海高原湖滨湿地退化定量评价模型:PC=0.548×PC1+0.291×PC2+0.159×PC3。并进行湿地退化评价:PC≤-0.300,原生湿地;-0.300 [1] Yang Y X. The 21st century hot point and forward position field of international wetland research from Quebec 2000-Millennium Wetland Event. Scientia Geogrophica Sinica, 2002, 22(2): 150- 155. [2] Yang Y X. Ecological environment deterioration, mire degeneration and their formation mechanism in the Zoige Plateau. Journal of Mountain Science, 1999, 17(4): 318- 323. [3] Frank D A. The interactive effects of grazing ungulates and aboveground production on grassland diversity. Oecologia, 2005, 143(4): 629- 634. [4] Boswell J S, Olyphant G A. Modeling the hydrologic response of groundwater dominated wetlands to transient boundary conditions: Implications for wetland restoration. Journal of Hydrology, 2007, 332: 467- 476. [5] Li K, Yang Y X, Yang Y, Han D Y. Characteristics and influence factors of the swamp degradation under the stress of grazing in the Zoige Plateau. Acta Ecologica Sinica, 2011, 31(20): 5956- 5969. [6] Manier D J, Hobbs N T. Large herbivores in sagebrush steppe ecosystems: livestock and wild ungulates influence structure and function. Oecologia, 2007, 152(4): 739- 750. [7] Prieto M R, Rojas F. Changing climatic and anthropogenic influences on the Bermejo wetland, through archival documents-Mendoza, Argentina, 16th—20th centuries. Clim. Past Discuss, 2011, (7): 3775- 3794. [8] Zamolyi A, Szekely B, Draganits E, Timar G. Neotectonic control on river sinuosity at the western margin of the Little Hungarian Plain. Geomorphology, 2010, 122(3/4): 231- 243. [9] Jiang J H, Huang Q, Sun Z D. Analysis of ecological environment of lake-wetland in Yangtze River basin. Ecology and Environment, 2006, 15(2): 424- 429. [10] Yang Y X, Yang Y, Liu C E. The basic academic issue of wetland and wetland science and ecological and environmental function of wetland ecosystem. Landscape Architecture China, 2009, 3: 14- 17. [11] Xiao D R, Tian K, Yang Y M, Yun H, Li N Y, Xu S G. Relationship between plant diversity and soil fertility in Napahai wetland of Northwestern Yunnan Plateau. Acta Ecologica Sinica, 2007, l6 (2): 523- 529. [12] Tian K. Mechanism and Process of Soil Degradation in Napahai Wetland on Yunnan Plateau. Changchun: Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences, 2004. [13] Tian K, Mo J F, Lu M, Chang F L, Yang Y X. Human disturbances on the ecological environment degradation of Napahai wetland in the upstream of Yangtze river. Resources and Environment in the Yangtze Basin, 2004, 13(3): 292- 295. [14] Wang Y, Li Q L, Guan L, Fang R, Jiang R. Effect of Traffic Noise around Napahai Wetland Highway on Birds. Chinese Journal of Zoology, 2011, 46(6): 65- 72. [15] Chinese SEPA. Water and wastewater analytical method (The fourth edition). Beijing: China Environmental Sciences Press, 2002. [16] The Institute of Soil Science, CAS. Analysis of Soil Physical and Chemical Characteristics. Shanghai: Shanghai scientific & Technical Publishers, 1987. [17] Ma K P, Huang J H, Yu S L, Chen L Z. Plant community diversity in Dongling mountain, Beijing, China Ⅱ: Species richness, evenness and species diversities. Acta Ecologica Sinica, 1995, 15(3): 268- 277. [18] Han D Y, Yang Y X, Yang Y, Li K. Species composition and succession of swamp vegetation along grazing gradients in the Zoige plateau, China. Acta Ecologica Sinica, 2011, 31(20): 5946-5955. [19] Zhang K, Tian K, Mo J F, Chang F L, Lu M, Li L Y. Impacts of hydrological periods on organic matters in Napahai wetlands. J. Lake Sci, 2007, 19(6): 705- 709. [20] Jin X C, Yan C Z, Xu Q J. The community features of aquatic plants and its influence factors of lakeside zone in the north of Lake Taihu. J. Lake Sci, 2007, 19(2): 151- 157. [21] Hu J M, Yuan H, Li J, Dong Y X, Ma B B. Variation of soil rapid available nitrogen components and its implication in Napahai lake-marsh region, Northwest Yunnan Mountain. Journal of Mountain Science, 2011, 29(3): 269- 275. [22] Venterink H O, Pieterse N M, Belgers J D, Wassen M J, DeRuiter P C. N, P, and K budgets along nutrient availability and productivity gradients in wetlands. Ecological Applications, 2002, 12(4): 1010- 1026. [23] Newman S, Grace J B, Koebel J W. Effects of nutrients and hydro period onTypha,Cladium, andElecocharis: Implications for everglades restoration. Ecological Application, 1996, 6(3): 774- 783. [24] Tian K, Chen B K, Bei R T, Luo K H, Wang Y W, Yang Y X. Application of in-situ technique to study N fluxes on degradaded soils. Acta Ecologica Sinica, 2003, 23(9): 1937- 1943. [25] Liu J P, Yang Q, Lu X G. Study on spatial distribution of soil nutrient elements of annular wetlands in Sanjiang Plain. Journal of Soil and Water Conservation, 2005, 19(2): 76- 79. [26] Yang Y X, Wang S Y. Human disturbances on mire and peat soils in the Zoige plateau. Resources Science, 2001, 23(2): 37- 41. [27] Andreas Grunig, ed. Mire and Man: Mire Conservation in a Densely Populated Country the Swiss Experience, Swiss Federal Institute for Forest, Snow and Landscape Research, Switzerland, 1994. 参考文献: [1] 杨永兴. 从魁北克2000-世纪湿地大事件活动看21世纪国际湿地科学研究的热点与前沿. 地理科学, 2002, 22(2): 150- 156. [2] 杨永兴. 若尔盖高原生态环境恶化与沼泽退化及其形成机制. 山地学报, 1999, 17(4): 318- 323. [5] 李珂, 杨永兴, 杨杨, 韩大勇. 放牧胁迫下若尔盖高原沼泽退化特征及其影响因子. 生态学报, 2011, 31(20): 5956- 5969. [9] 姜加虎, 黄群, 孙占东. 长江流域湖泊湿地生态环境状况分析. 生态环境, 2006, 15(2): 424- 429. [10] 杨永兴, 杨杨, 刘长娥. 湿地与湿地科学基本理论问题与湿地生态系统的生态、环境功能. 景观设计学, 2009, 3: 14- 17. [11] 肖德荣, 田昆, 杨宇明, 袁华, 李宁云, 徐守国. 高原退化湿地纳帕海植物多样性格局特征及其驱动力. 生态环境, 2007, l6(2): 523- 529. [12] 田昆. 云南纳帕海高原湿地土壤退化过程及驱动机制. 北京: 中国科学院研究生院, 2004. [13] 田昆, 莫剑锋, 陆梅, 常凤来, 杨永兴. 人为活动干扰对纳帕海湿地环境影响的研究. 长江流域资源与环境, 2004, 13(3): 292- 295. [14] 王云, 李麒麟, 关磊, 房锐, 姜睿. 纳帕海环湖公路交通噪声对鸟类的影响. 动物学杂志, 2011, 46(6): 65- 72. [15] 国家环境保护总局. 水和废水监测分析方法(第四版). 中国环境科学出版社, 2002. [16] 中国科学院南京土壤所. 土壤理化性质分析. 上海: 上海科技出版社, 1987. [17] 马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究 Ⅱ:丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268- 277. [18] 韩大勇, 杨永兴, 杨杨, 李珂. 放牧干扰下若尔盖高原沼泽湿地植被种类组成及演替模式. 生态学报, 2011, 31(20): 5946-5955. [19] 张昆, 田昆, 莫剑锋, 常凤来, 陆梅, 李宁云. 水文周期对纳帕海高原湿地草甸土壤碳素的影响. 湖泊科学, 2007, 19(6): 705- 709. [20] 金相灿, 颜昌宙, 许秋瑾. 太湖北岸湖滨带观测场水生植物群落特征及其影响因素分析. 湖泊科学, 2007, 19(2): 151- 157. [21] 胡金明, 袁寒, 李杰, 董云霞, 马彬斌. 滇西北纳帕海湿地区域土壤速效氮组分的分异及指示意义. 山地学报, 2011, 29(3): 269- 275. [24] 田昆, 陈宝昆, 贝荣塔, 罗开华, 王有位, 杨永兴. In- situ方法在研究退化土壤氮库时空变化中的应用.生态学报, 2003, 23(9): 1937- 1943 [25] 刘吉平, 杨青, 吕宪国. 三江平原典型环型湿地土壤营养元素的空间分异规律. 水土保持学报, 2005, 19(2): 76- 79. [26] 杨永兴, 王世岩. 人类活动干扰对若尔盖高原沼泽土、泥炭土资源影响的研究. 资源科学, 2001, 23(2): 37- 41. ProcessanalysisandevaluationofwetlandsdegradationbasedonPCAinthelakesideofNapahai,NorthwestYunnanPlateau SHANG Wen, YANG Yongxing*, HAN Dayong KeyLaboratoryofYangtzeRiverWaterEnvironment,MinistryofEducation,StateKeyLaboratoryofPollutionControlandResourcesReuse,CollegeofEnvironmentalScienceandEngineering,TongjiUniversity,Shanghai200092,China Napahai wetland is a unique and seasonal marsh wetland located in the middle of Hengduan Mountains, northwestern Yunnan Plateau. In the recent decades, it has been seriously degraded under human disturbances, mainly including drainage and overgrazing. In the present paper, two kinds of typical degradation transects were established to evaluate the synergistic impacts of overgrazing stress and water regime and the single impacts of overgrazing stress in the lakeside of Napahai. The two transects were further classified into four degradation gradients, including primary wetland(PW), lightly degraded wetland (LW), moderately degraded wetland (MW) and severely degraded wetland (SW). Cluster analysis and principal component analysis (PCA) were conducted to explore the process and mechanism of wetland degradation as well as to evaluate the degree of wetland degradation quantitatively at system level of “vegetation-water-soil”. The results were as follows: the numbers of plant families, genera and species as well as species diversity and richness all increased gradually from PW to SW, whereas species evenness remained stable. The plant community succession is in the direction of aquatic plant community →swamp plant community →swamp meadow plant community →meadow plant community. Correspondingly, the community changed towards more complicated structure characterized by the decreasing of helophyte and increasing of hygrophyte and mesophyte. Overgrazing stress changed the inherent mode of plant community succession and accelerated the degraded stage of “swamp meadow community” to some extent, although the direction of succession had not changed. The degradation characteristics of water properties varied in different disturbance type, the water indices of salinity, total alkalinity, total hardness, total nitrogen, ammonia nitrogen, nitrate nitrogen and total phosphorus all increased along the GD gradients, whereas decreased along the GAD gradients, depending on the water regime. Therefore, hydrologic alteration had a significant effect on wetland water properties. The changes of wetland soil properties were similar in the two disturbances. Among them, the contents of organic matter, total nitrogen and available nitrogen decreased, total phosphorous and total potassium (K) increased, whereas the available phosphorous and available potassium (K) showed no obvious changes. The vertical distribution for each nutrient content in the soil profiles of degraded wetlands varied greatly. Soil total phosphorous, available phosphorous and available potassium (K) decreased along the soil depth gradients. It showed that soil nitrogen content was the most obvious characteristics of wetlands degradation. In order to identify the driven factors of wetlands degradation under the multi-factorial stresses, PCA was employed to synthesize the degradation information from multiple aspects as discussed in this paper. Thirty indices based on vegetation, water and soil were simplified into three principal components by PCA, explained 54.8%, 29.1% and 15.9% of the total variance, respectively. PCA results indicated that plant community was the most representative of wetlands degradation, and the soil structures and nutrient contents changed significantly along the degradation gradients of lakeside wetlands, Napahai. By contrast, the contributions made by water nutritional contents were lowest compared with vegetation and soil for wetlands degradation. Based on the three principle components derived from PCA, a multiple linear regression model, PC = 0.548 × PC1 + 0.291 × PC2 + 0.159 × PC3, was conducted to calculate the scores for each degradation stage, which could be taken to evaluate the wetlands degradation. degradation; process; PCA; evaluation; Napahai wetlands 国家自然科学基金资助项目( 41171074, U0933601, 40771013) 2012- 05- 06; 2012- 11- 19 *通讯作者Corresponding author.E-mail: wetlands@tongji.edu.cn 10.5846/stxb201205060653 尚文, 杨永兴, 韩大勇.基于PCA的滇西北高原纳帕海湿地退化过程分析及其评价.生态学报,2013,33(15):4776- 4789. Shang W, Yang Y X, Han D Y.Process analysis and evaluation of wetlands degradation based on PCA in the lakeside of Napahai, Northwest Yunnan Plateau.Acta Ecologica Sinica,2013,33(15):4776- 4789.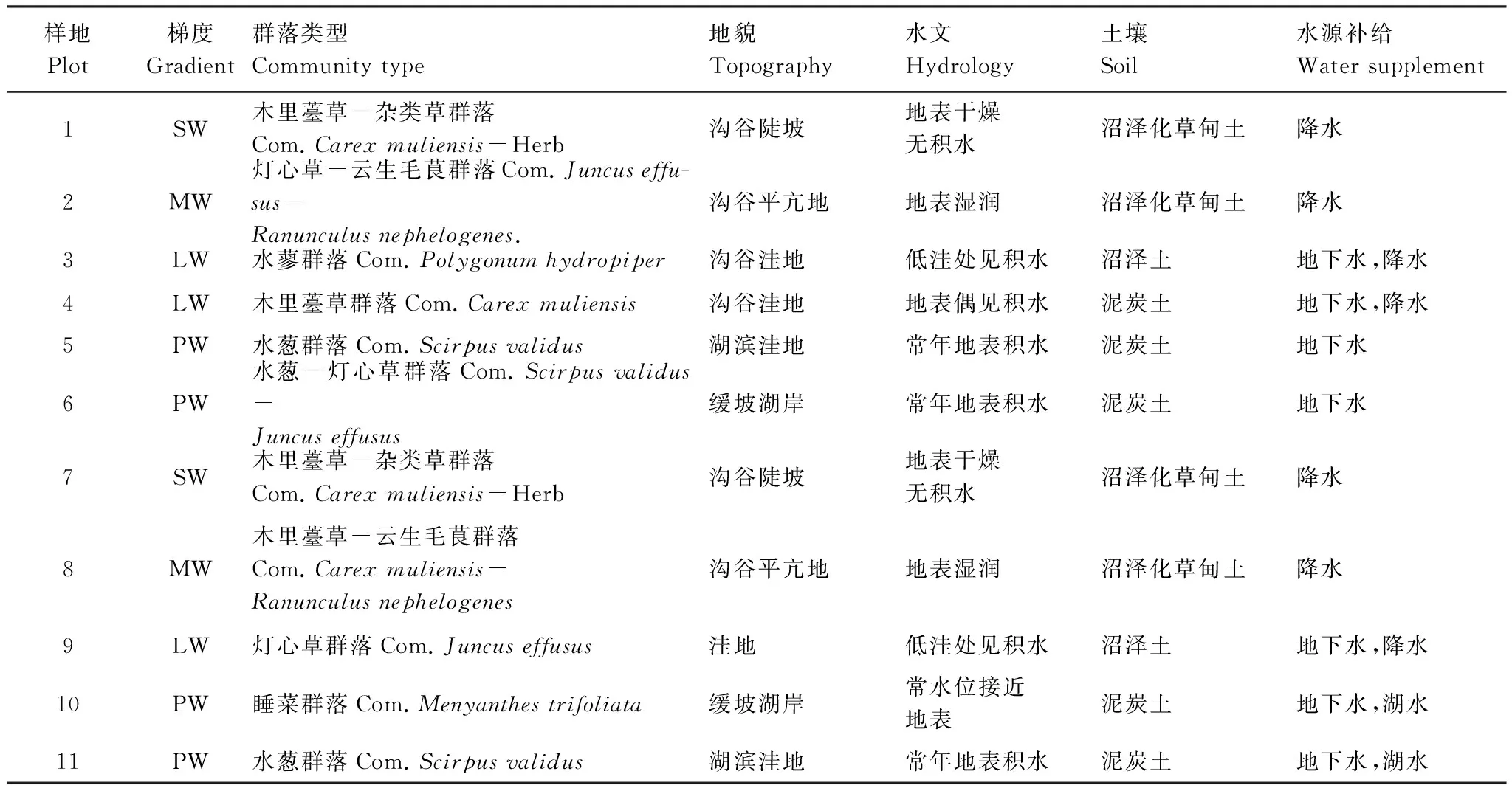
3 湿地退化评价
4 讨论
5 结论
