花臭蛙线粒体DNA 12S及16SrRNA序列的测定
2013-12-04王晶晶纪会会
王晶晶, 纪会会, 焦 山
(合肥师范学院 生命科学系,安徽 合肥230601)
1 引言
动物线粒体DNA呈共价闭环双链结构,在一级结构上,其进化速率为核DNA的5~10倍,绝大多数都表现为母系遗传。其基因组结构相对稳定、简单、无组织特异性、便于分析,而且容易纯化得到,使其成为研究动物起源进化及群体遗传分析的理想的研究对象[1-2]。碱基序列分析虽然技术要求高,花费大,但通过测定线粒体DNA全序列或部分基因片段序列可以了解碱基的置换、缺失、插入等实际情况,并可比较不同物种或个体碱基相关序列的差异,从而探讨进化关系,因此比其他分析方法具有很大的优点,目前已被应用到分类进化领域研究中[3-4]。
M.和S.Nass发现线粒体DNA(mtDNA)后,又发现线粒体DNA还能合成12S和16SrRNA及22种tRNA,随着分子系统学以及DNA分子技术的发展,从DNA水平研究物种间亲缘关系以及进行物种辅助鉴定已经逐渐成为一种新的手段[3-6]。已知动物线粒体DNA(mtDNA)具有分子量小、母性遗传以及比核基因DNA进化速度快等特点,其中的12S和16SrRNA片段更是保守,所以Hedges and Maxsom (1993)和 Hay et al.(1995)便提出了线粒体12S和16SrRNA基因可以用于研究两栖动物系统发育[7],不少学者做了大量的工作。现已被广泛应用于种内和种间系统发育研究中,尤其在蛙科的系统发育方面[7-8]。
两栖动物是从水生到陆生进化上的一个关键环节,在系统进化上具有承前启后的重要作用[9]。两栖类是脊椎动物进化从水生到陆生重大转变过程中的一个过渡类群,在地球生命进化研究中占有重要地位。花臭蛙(Rana schmackeri)隶属两栖纲(Amphibia)、无尾目(Anura)、蛙科(Ranidae),鼓膜大,约为第三指吸盘的2倍[10];上眼睑、体后背部及后肢背面均无小白刺,体侧无背侧褶;指、趾具吸盘,纵径大于横径,均有腹侧沟;体背面为绿色,间以棕褐色或褐黑色大斑点,多近圆形并镶以浅色边[9-10];分布于江苏、浙江、安徽、福建、江西、河南、湖北、湖南、广东、广西、四川、贵州、陕西、甘肃等地,多见于较开阔的山溪及附近潮湿处以及常蹲在有苔藓的岩石上,其生存的海拔范围为200至1500米,是中国的特有物种[10]。
在具有独特进化地位的两栖类动物中,对系统进化上的保守性状进行深入研究,将有助于花臭蛙的系统分化与分类问题提供依据[11-13]。本实验是通过测定花臭蛙线粒体12S和16SrRNA基因序列,再利用ClustalX软件分析与其他相关物种(例如:黑斑蚊、花臭蛙、天台蛙)的同源关系,为将来花臭蛙的分类提供分子生物学方面的证据。
2 材料和方法
2.1 材料
实验所用的花臭蛙于2011年6月采自安徽黄山自然保护区海拔500米附近的山溪,置-20℃冰箱保存备用。
实验主要试剂:购买自TIANGEN的TIANa-mp Genomic DNA Kit、Ex Taq 酶、dNTPs、1000bpLadder、10×Ex Taq缓冲液。
2.2 方法
2.2.1 基因组DNA的提取
取0.1g花臭蛙肌肉组织,利用TIANGEN公司的TIANamp Genomic DNA Kit提取其基因组DNA。将乙醇沉淀后DNA溶解于50μL灭菌蒸馏水,测定DNA浓度后于4℃存放。为防止DNA污染,在基因组DNA提取过程中,以阴性作为对照,用TE溶液代替溶解样品。为避免交替污染,DNA提取分3次在不同的时间进行。
2.2.2 引物设计及PCR扩增
根据从GenBank检索到的其它相关蛙科物种的12S和16SrRNA基因序列,分别设计花臭蛙12S和16SrRNA基因引物:
12SrRNA——F 5′-GAACTACGAGCC-3′,12SrRNA——R 5′-GTGTACGCGTCC-3′;
16SrRNA—F 5′-UGAGCAUAGUUG — 3′,16SrRNA—R 5′-AUAGGAUCAC-3′。在PCR仪(Backman)上进行PCR扩增。PCR反应总体积为25μL,包括:浓度20~50μmol/μL引物各1μL,10×Ex Taq 缓冲液(TIANGEN)2.5μL,dNTPs(TIANGEN)2μL,1Lg模板 DNA 1μL,Ex Taq 酶(TIANGEN)1.25U,其它为灭菌蒸馏水。PCR反应程序如下:12SrRNA(94℃预变性300s,然后94℃变性45s,45℃退火45s,72℃延伸45s,35个循环,最后72℃延伸420s);16SrRNA(95℃预变性5min,95℃变性30 s,57℃退火30s,72℃延伸30s,循环次数为30。循环结束后再72℃延伸10min)。用1.5% 琼脂糖凝胶电泳检测PCR扩增片段大小,溴化乙锭染色,紫外透射仪观察并照相记录。
2.2.3 序列测定
观察到条带后将样品送至上海生工进行花臭蛙12S和16SrRNA基因片段的序列测定。
2.2.4 数据分析
利用DNASTAR软件对测得的12S和16S rRNA基因序列进行编辑排序和计算,再利用ClustalX软件分析其与黑斑蛙(Rana nigromaculata,GenBank登陆号:JN541318.1)、天台蛙(Rana tientaiensis,GenBank登陆号:AF205560.1)、大壁虎(Gekko gecko,GenBank登陆号:DQ093192)、卡氏小鼠 (Mus caroli,GenBank 登陆号:JQ287760.1)、大猩猩(Gorilla gorilla,GenBank登陆号:AJ627520.1)以及人(Homo sapiens,Gen-Bank登陆号:AF121220.1)的同源关系。
3 结果与分析
3.1 花臭蛙线粒体12S和16SrRNA基因片段的PCR扩增
利用GenBank检索到的其它相关蛙科物种的12S与16SrRNA基因序列设计的简并引物进行花臭蛙线粒体基因12S和16SrRNA片段的PCR扩增,扩增产物用琼脂糖凝胶进行电泳分离,可见清晰的特异性扩增条带,长度与预期一致,而且空白对照试验未出现扩增产物。(图1是扩增结果)
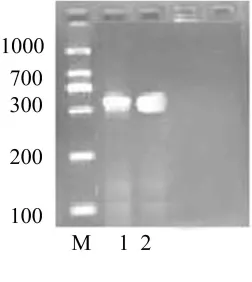
图1 花臭蛙线粒体12S与16SrRNA基因片段PCR扩增结果
3.2 花臭蛙线粒体12S和16SrRNA基因片段序列的测定
测定出花臭蛙(Rana schmackeri)线粒体基因12SrRNA 347bp的基因片段(如下图2),其中A、T、G 、C 的含量分别是 105bp(30.26%),78bp(22.48%),76bp(21.90%),88bp(25.36%)。A+T含量为52.74%,C+G含量为47.26%。

图2 检测得到的花臭蛙的线粒体12SrRNA基因序列
花臭蛙16SrRNA序列长度为380bp(如下图3),对测序结果统计4种碱基(A、G、C、T)在核苷酸序列中的平均含量,发现A、G、C、T含量分别为:31.8% (121 个)、20.8% (79 个)、25.8% (98个)、21.6%(82个);其中 G+C为47%,A+T为53%。

图3 检测得到的花臭蛙的线粒体16SrRNA基因序列
3.3 线粒体12S和16SrRNA基因序列碱基组成的比较
根据现有的研究,通过测定动物的线粒体基因全序列或者部分序列,再相比较物种或个体碱基序列差异可以研究探进化关系。在GenBank中下载了大绿蛙(Rana livida)、黑斑蛙(Rana nigromaculata)、天 台 蛙 (Rana tientaiensis)、林 蛙 (Rana chensinensis)及大蹼林蟾(Bombina maxima)的线粒体12S和16SrRNA同源序列进行进行分析比对(见表1、表2)。
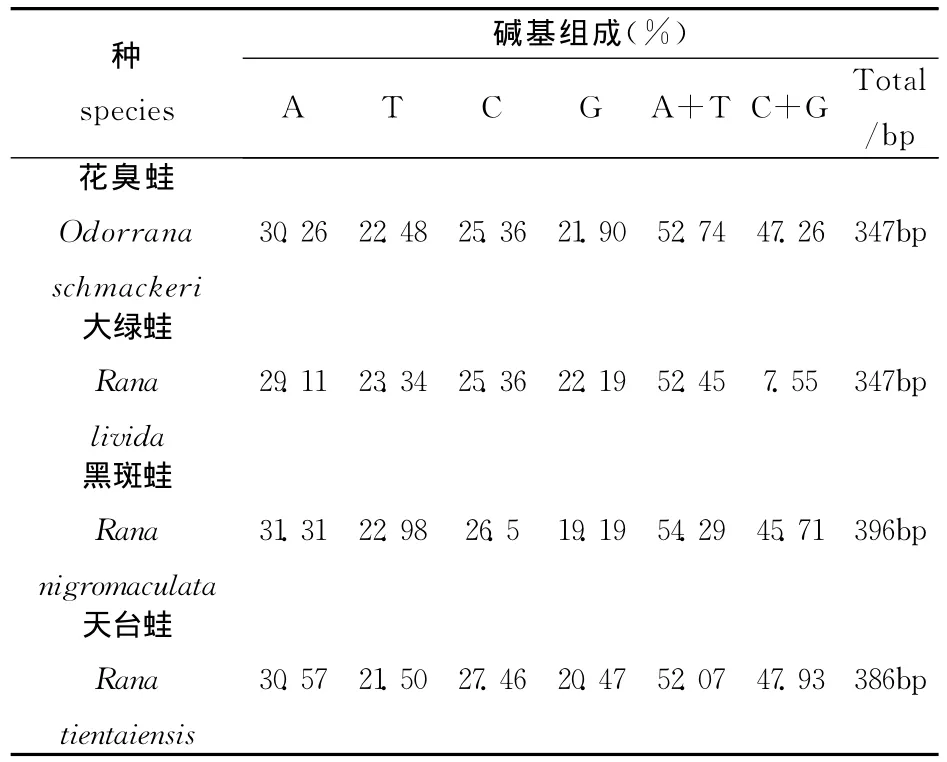
表1 四种蛙的12SrRNA基因序列的碱基构成情况
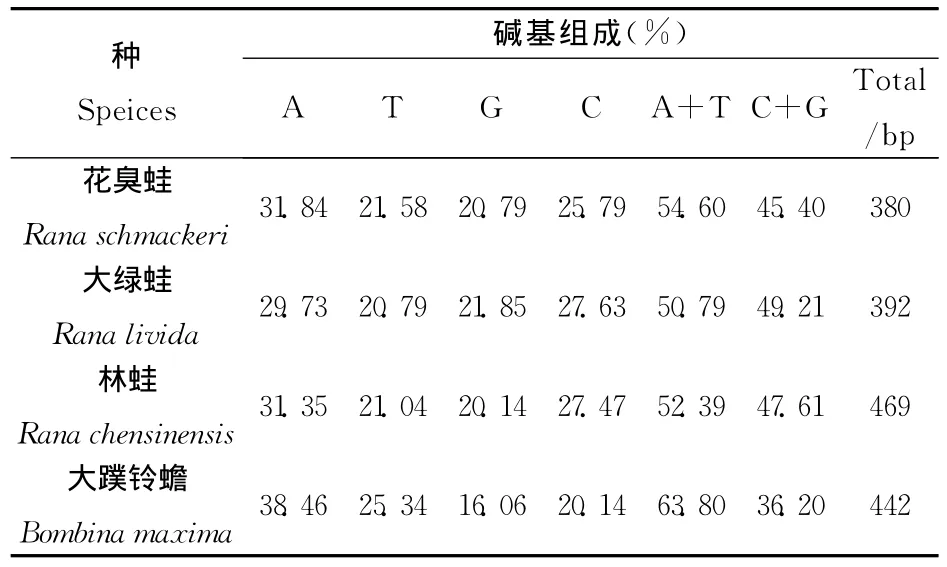
表2 四种两栖类物种16SrRNA基因片段碱基组成
在表1中,大绿蛙、黑斑蛙和天台蛙的序列长度分别为347bp、350bp、386bp,其 A、T、C、G 的含量分 别 为 29.11%、23.34%、25.36%、22.19%;31.31%、22.98%、26.52%、19.19%;30.57%、21.50%、27.46%、20.47%。经试验测定得到的花臭蛙347bp的序列中 A、T、C、G的含量分别为30.26%、22.48%、25.36%、21.90%。与上面三种蛙的线粒体12SrRNA序列对比之后不难发现,花臭蛙、大绿蛙、黑斑蛙和天台蛙四种种蛙中A+T的含量平均值十分相近,在52.07%~54.29%之间,略高于C+G的含量(47.26%~47.93%)。
同理,将花臭蛙16SrRNA的序列与大绿蛙、林蛙及大蹼铃蟾的序列进行分析(见表2),经计算各个碱基在相应的基因片段所占比例相似,且A+T的含量也都大于G+C的含量。直接反映出了以上四个物种具有较高同源性,也表明16SrRNA在蛙科进化中的保守性。
3.4 花臭蛙与其它物种的同源性比较
利用ClustalX软件,从GENBANK下载一些物种的相关序列,分别对花臭蛙的12S和16S进行序列的同源性分析(见表3、表4),可得到花臭蛙线粒体12SrRNA与天台蛙的相关序列同源性为80.45%,与鱼类中的黯纹东方鲀相关序列的同源性63.25%,与爬行类四线棉蛇的相关序列的部分片段同源性为65.43%,与哺乳类家鼠相关基因的同源性为66.34%。
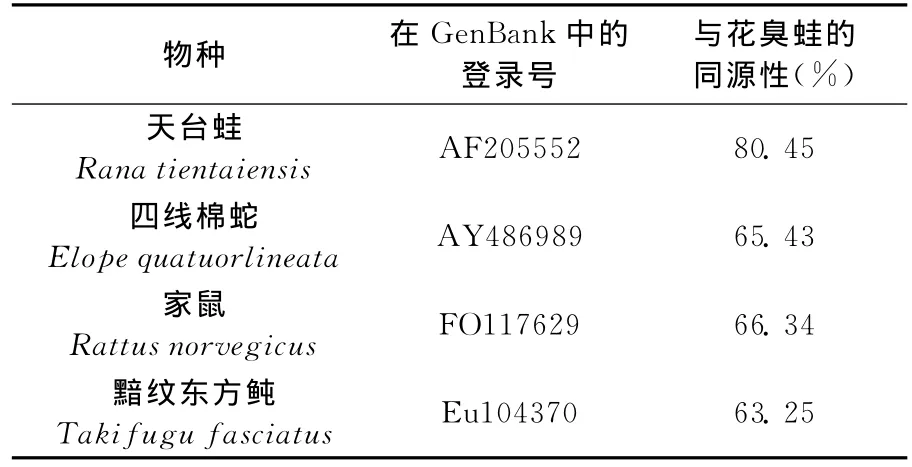
表3 花臭蛙与其他物种12SrRNA基因的同源性比较
花臭蛙16SrRNA基因与同为两栖类蛙属的天台蛙(Rana tientaiensis)同源性高达86%,与爬行类的大壁虎(Gekko gecko)同源性为74%,与四线棉蛇(Elope quatuorlineata)的同源性为78%,与鱼类的斑马鱼(Danio rerio)同源性为68%,与哺乳类家鼠的同源性为65%。
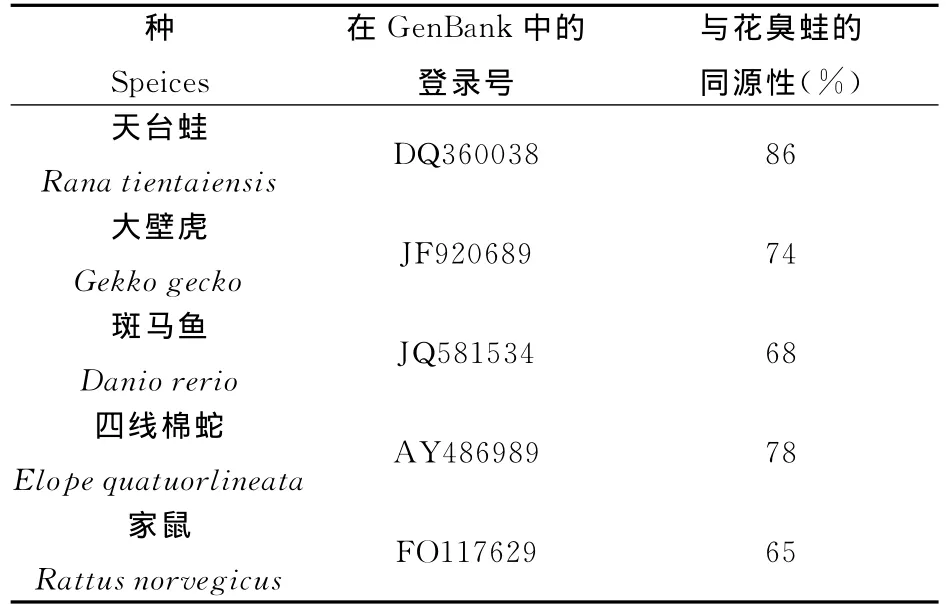
表4 花臭蛙与其他物种16SrRNA基因的同源性比较
4 讨论
4.1 用线粒体基因研究的意义
线粒体基因能够独立地进行复制和转录,且分子量小,结构较简单,使得其更容易从组织中分离、提纯;线粒体基因mtDNA因其母系遗传、无内含子、缺少重组的遗传物质,而常被用于物种的系统进化和分类地位的研究[13-14]。其中,16SrRNA 基因进化程度适中,通过碱基序列分析,再与其它相关物种进行同源性分析,比较差异,来探讨进化关系,其常作为分子标记用于哺乳动物、鸟、两栖、爬行类等物种系统进化的研究[15]。
4.2 花臭蛙与其它物种12S和16SrRNA的同源性分析
利用ClustalX软件将花臭蛙与相关物种的目的片段进行同源性分析,结果显示:花臭蛙与天台蛙的同源性最高,达到了(12SrRNA达到80.45%,16SrRNA达到86%),而与爬行类的四线棉蛇等相关物种具有较高的同源性(12SrRNA达到65.43%,16SrRNA达到65%),与哺乳类的家鼠、鱼类的斑马鱼具有一定的同源性(与家鼠的12S rRNA达到66.34%,16SrRNA达到65%),这与两栖类内部亲缘关系较近,两栖类与其它相关物种的亲缘关系较远相符合,符合物种的进化规律,下一步可通过构建系统进化树来研究花臭蛙在蛙科分类中的关系和地位[16-18]。
4.3 展望
随着DNA的序列分析、PCR技术的应用,mtDNA在动物起源、种群分化等方面已经取得了具有应用价值的成果。mtDNA的碱基序列分析虽然是个浩大的工程,不仅花费很高,技术要求也很高,但mtDNA进化速度适中,基因高度保守,通过测定物种个体mtDNA的碱基序列,然后与其它相关物种的碱基序列进行同源性分析,比较差异,来探讨物种的进化地位,这相对于其它分析方法,具有很大的优势[1-4]。
目前在我国,两栖动物的生存现状不容乐观,部分甚至有濒临灭绝的危险。我们可通过分子生物学方面的手段来研究动物线粒体基因,了解物种的生物学特性,从而制定出相应的保护措施,缓解物种的灭绝趋势[8-11]。
[1]Clary D O and D R Wolstenholme.The mitochondrial DNA molecular sequence of Drosophila yakuba,gene organization and genetic code[J].Mol.Evol,1985,22:252-271.
[2]郑冬,刘学东,马建章.12SrRNA基因及其二级结构在系统学研究中的应用[J].东北林业大学 学报,2003,31(3):59-61.
[3]根井正利,苏德海尔 库马.分子进化与系统发育[M].北京:高等教育出版社,2002,21(2):39-41.
[4]毕相东,侯林,刘晓惠,等.核糖体rRNA基因在海洋动物分子系统学中的应用[J].应用与环境生物学报学报,2005,11(6):779-783.
[5]Brown W M.Evolution of animal mitochondrial DNAs,M Nei,Koehned R K.Evolution of genes and proteins[M].Sunderland MA:Sinauer,1983,62-88.
[6]Hedges S B and Maxsom L R.A molecular perspective on lissamphibian phylogeny[M].Herpetol.Monnogr,1993,7:27-42.
[7]Matsui M,Shimada T,Ota H,et al.Multiple invasions of the Ryukyu Archipelago by Oriental frogs of the subgenus Odorrana with phylogenetic reassessment of the related subgenera of the genus Rana[J].Mol.Phylogenet.Evol,2005,37(3):733-742.
[8]赵尔宓,张学文,赵蕙,鹰岩,中国两栖纲和爬行纲动物校正名录[M].2000,19(3):196-2070.
[9]李成,叶昌媛,费梁,我国臭娃属物种的系统发育研究,中国动物科学研究[M].北京:中国林业出版社,1990,1194-1195.
[10]FEILiang,YE ChangYuan LI Cheng.Descriptions of two new species od the genus Odorrana in china(Anura:Randae)[J].动物学分类报,2001,26(1):108-114.
[11]江建平,谢锋,金义文等,基于12S和16SrRNA序列的濡蛙属部分物种的系统发育关系[J].动物学研究,2005,26(1):61-68.
[12]王广力,何舜平,黄松,等.美姑脊蛇Achalinus meiguensis线粒体基因组全序列及系统发育地位研究[J].科学通报,2009,54(9):1250-1261.
[13]Kocher W.K,Thomas A,Meyer S.V,et al.Dynamics of mitochondrial DNA evolution in animals:Amplifitication and sequencing with conserved primers[J].Proc Natl Acad Sci USA,1989,86:6196-6200.
[14]江建平,周开亚,从12SrRNA基因序列研究中国24种蛙的进化关系[J].动物学报,2001,47(1):38-44.
[15]叶昌媛,费梁,松井正文,我国日本林蛙的分类研究[J].两栖爬行动物学研究,1995,1-87.
[16]Matsui,M.,A taxonomic study of the Rana narina complex,with description of three new species[J].Zool.J.Linn.Soc.1994,111:385-415.
[17]Boulenger.G.A.A Monograph of the south Asian,Malanesian and Australian Frogs of the Genus Rana.Rec[J].Ind Mus.1920,20:1-226.
