中国白兔IL-6基因的克隆与序列分析
2013-11-22张夏兰王红宁张安云
张夏兰,陈 军,王红宁,杨 鑫,张安云
(1.四川农业大学动物科学院,四川 雅安625014;2.四川大学生命科学学院动物疫病防控与食品安全四川省重点实验室,四川 成都610064;3.重庆市巴南区动物疫病预防控制中心,重庆 巴南401320;4.重庆市动物卫生监督所,重庆 巴南401147)
白介素6是已知的功能最广泛的细胞因子,作为一种重要的免疫调节剂,在细胞免疫中有广谱性,在动物疾病的预防、治疗和构建基因工程疫苗中显示出广阔的应用前景,在家兔上做白介素6的研究实属必要。本试验基于IL-6在免疫学方面特有的生物学活性,根据GenBank上登录的兔IL-6的基因序列,结合人IL-6的基因序列,设计引物,从中国白兔的淋巴细胞RNA中扩增出IL-6基因。本试验在国内尚属首次,具有一定学术价值和应用前景。
1 材料与方法
1.1 材料 1~2月龄的中国白兔,购自四川某肉兔场。RNA抽提试剂盒、反转录酶AMV、Oligo(dt)、RNA 酶 A、Taq酶、d NTP 、EcoRⅠ、HindⅢ、RNA酶抑制剂、T连接酶、质粒抽提试剂盒、载体pMD18-T、DNA Marker DL-2 000,均购自宝生物工程(大连)有限公司;ConA,购自Sigma公司;营养液1640,购自天泰公司,兔淋巴细胞分离液,购自天津市灏泽生物制品科技有限公司。
1.2 方法
1.2.1 兔淋巴细胞的分离及总RNA的提取 无菌采取兔外周血,肝素抗凝,在无菌条件下加入等量的PBS稀释兔血,用兔淋巴细胞分离液分离兔PB-MC.兔淋巴细胞总RNA的提取是按照Bio Spin RNA抽提试剂盒操作说明书进行。
1.2.2 引物的设计与合成 根据GenBank上登录的兔的白介素6序列(序列号:AF169176),设计1对引物,在引物的两侧分别加上酶切位点HindⅢ和EcoRⅠ,并在没酶切位点上添加保护碱基。引物序列如下:
上 游: 5′-AGCAAGCTTATGAACTCCTTCACAAGCG-3′;下游:3′-GGACAGGTAACCTGTGTATTCTTAAGTAA-5′。引物由宝生物工程(大连)有限公司合成,预期产物片段长度约为726bp。
1.2.3 中国白兔IL-6基因RT-PCR 扩增 30μL的RT反应体系:总 RNA 9μL,10mmol/L dNTP 2μL,Oligo dT 1μL,RNAsin 0.5μL混匀,70℃4 min,再加入5×RT Buffer 4μL,Mg2+3μL,反转录酶AMV 0.5μL,RNAsin 0.5μL,混匀后按如下程序进行 RT反应:30℃10min,42℃45min,95℃5min。反应产物进行PCR反应。PCR的反应体系为50μL:10×PCR Buffer 5μL,Mg2+3μL,10mm d NTP 1μL,上游引物(30pmol/L)1μL,下游引物(30pmol/L)1μL,反转录产物c DNA 1 μL,TaqDNA聚合酶0.3μL,H2O 补至50μL,混匀,95℃预变性5min后按以下条件进行PCR反应:94℃30s,54℃50s,72℃60s,35个循环之后,72℃延伸10min,4℃10min后结束反应。取5μL反应产物点样于1%琼脂糖凝胶电泳,80V电泳30 min,在凝胶成像系统上观察结果并拍照。
1.2.4 中国白兔IL-6的克隆及鉴定 PCR产物用1%琼脂糖凝胶电泳,参照Marker DL-2 000切取并回收目的条带,按常规方法将目的片段连入载体pMD18-T,转化JM109宿主菌,用菌落PCR、重组质粒酶切及重组质粒PCR筛选阳性质粒。将阳性质粒送往宝生物工程(大连)有限公司进行序列测定。
1.2.5 序列分析 应用DNASRAR软件,将测得序列,与GenBank上登载的世界其他5个品种的兔IL-6的核苷酸序列比较,探讨中国白兔IL-6的变异规律。
2 结果
2.1 兔IL-6基因的RT-PCR扩增结果 从ConA诱导的兔外周血淋巴细胞中分离RNA后,应用RT-PCR方法扩增得到的约750bp的条带,其大小与上下游引物涵盖的兔IL-6核酸序列长度一致(图1)。
2.2 兔IL-6基因的克隆与鉴定 将PCR纯化产物与pMD18-T载体连接后,并转化E.coliJM109感受态细胞,在37℃培养箱培养12h后,用转化子作菌落PCR鉴定,结果扩增出约750bp大小的一条带(见图1)。
菌落PCR阳性转化子质粒,用限制性内切酶HindⅢ和EcoRⅠ酶切鉴定,结果具有与目的片段大小一致的电泳带(见图1),表明RT-PCR产物成功地亚克隆到pMD18-T载体上。

图1 兔IL-6RT-PCR及质粒pMD-IL-6鉴定
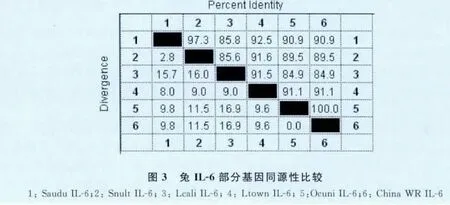
2.3 序列分析 经序列测定,扩增出的基因片段长726bp(见图2),共编码241个氨基酸,与欧洲兔(Oryctolagus cuniculus)IL-6基因(AF169176)同源性为100%,从而证明了本研究成功的克隆出了中国白兔IL-6基因。中国白兔IL-6与GenBank上登录的白尾野兔、高山白尾野兔、加利福利亚黑尾长腿野兔和白尾长腿野兔的IL-6的基因序列同源性比较结果显示,其种间的同源性为84.9%~91%,见图2。进化树分析结果表明,本研究所用的中国白兔与欧洲兔的亲源性最近,与加利福利亚黑尾长腿野兔的亲源性最远。详细情况见图3。
远端序列比较发现,相对于其他4种兔的IL-6基因序列,欧洲兔和中国白兔在通常的终止子的地方有特征性的突变,导致欧洲兔和中国白兔IL-6基因序列终止密码子位于延长81bp的下游区。


3 讨论
白介素6是一种多功能细胞因子,研究表明,白介素6具有疫苗分子佐剂和抗生素替代物的潜能。中国白兔IL-6基因序列,与欧洲兔IL-6的基因序列相同,相对于GenBank上登录的其他四种野兔的IL-6基因,其同源性差异较大。由于在通常的终止子的地方有变异,下一个终止密码子位于延长81 bp的下游区,故中国白兔IL-6基因序列3'端的编码区有27个氨基酸的延长。研究发现[1],家兔和野兔这两个谱系完全不同,家兔来自于混合家兔谱系实验兔,野兔是通过野外铺捉得来的,而这些野兔是一百多年前,最初的欧洲兔由于生活史的扩散,释放到澳大利亚。两种棉尾兔属和两种野兔属,都来自北美而不是欧洲,故其IL-6基因在通常的位置出现了常见的终止子。因此,IL-6在中国白兔和欧洲兔上突变的终止子具有种的特异性,可推断中国白兔和欧洲白兔有很近的亲缘关系,可能来自相同的谱系。
兔IL-6蛋白质羧基端延伸27个氨基酸的影响不得而知。学者们提出六步格的白介素6受体复合物的模式,这个复合物由2个三聚物组成,包含IL-6,IL-6R,和gp 130单个分子[2-5],指出IL-6分子的氨基和羧基末端位于复合物的外缘。虽然接近羧基端的几个残基对结合IL-6受体是决定性的[6-7],但是一个延伸的羧基端将干扰这些相互作用是不太可能的。研究显示[8-9],鼠IL-6 羧基端3个和5个残基的小延长并未影响其生物学活性。
[1] Chapman J A,Flux J E C.Introduction and overview of the lagomorphs in rabbits,hares and pikas[J].Status Survey and Conservation Action Plan,IUCN,Gland,Switzerland,2012,1-6.
[2] Ward L D,Howlett G J,Discolo G,etal.High affinity interleukin-6receptor is a hexameric complex consisting of two molecules each of interleukin-6,interleukin-6receptor,and gp-130[J].Biol Chem ,1994,269:23286-23289.
[3] Simpson R J,Hammacher A,Smith D K,etal.Interleukin-6 structure-function relationships[J].Protein Sci,1997,6:929-955.
[4] Somers W,Stahl M,Seehra J S.A crystal structure of interleukin 6:implications for a novel mode of receptor dimerization and signalling[J].EMBO ,1997,16:989-997.
[5] Fontaine V,Savino R,Arcone R,etal.Involvement of the Arg179in the active site of human IL-6[J].Eur Biochem ,1993,211:749-755.
[6] Savino R,Lahm A,Giorgio M,etal.Saturation mutagenesis of the human interleukin6receptor binding site:implications for its three dimensional structure[J].Proc Natl Acad Sci USA ,1993,90:4067-4071.
[7] Neipel F,Albrecht J C,Ensser A,etal.Human herpesvirus 8encodes a homolog of interleukin-6[J].Virol,1997,71:839-842.
[8] Burger R,Neipel F,Fleckenstein B,etal.Human herpesvirus type 8interleukin-6homologue is functionally active on human myeloma cells[J].Blood,1998,91:1858-1863.
[9] Barouch D H,Santra,Steenbeke T D,etal.Augmentation and suppression of immune responses to an HIV-1DNA vaccine by plasmid cytokine/Ig administration[J].Journal of Immunology(Baltimore),1998,161:1875-1882.
