产胸苷磷酸化酶短乳杆菌融合菌株的构建及其特性*
2013-11-19王伟洁李红梅薛芳高露娇黄艳青
王伟洁,李红梅,薛芳,高露娇,黄艳青
1(上海理工大学医疗器械与食品学院,上海,200093)
2(中国水产科学研究院东海水产研究所,上海,200090)
核苷磷酸化酶分3类:嘌呤核苷磷酸化酶、尿苷磷酸化酶和胸苷磷酸化酶[1],其中胸苷磷酸化酶主要用来合成核苷类药物,在“补救途径”中催化脱氧胸苷可逆磷酸化反应,提供2-脱氧核糖-1-磷酸,释放碱基胸腺嘧啶,加入另一种碱基生成新的核苷,在肿瘤治疗中有着重要的作用[2-3]。目前国内外报道的核苷磷酸化酶的产生菌主要有大肠杆菌、产气肠杆菌、乙酰短杆菌以及欧文杆菌等[4],短乳杆菌产核苷磷酸化酶却鲜有报道。
目前酶法合成核苷类物质的酶多数来源于野生菌株,基因工程菌尽管能有效提高酶产量,但构建步骤繁琐,成本较高,仅有少量的报道。文献调研表明,提高野生菌株产酶能力的方法主要集中在产酶发酵培养基的优化、传统诱变育种及基因组重排等技术领域。大量的研究结果表明培养基优化对菌体产酶量提高通常不明显,传统诱变技术具有盲目性和两面性,诱变育种时间长等缺点。在原生质体融合技术基础上发展起来的基因组重排技术不需预先掌握微生物的遗传,只需在了解微生物遗传性状的基础上实现对微生物的定向育种,此方法不但简化了选育步骤,缩短了育种周期,而且为经过多年诱变对理化诱变已不敏感的菌株提供新的改良方法,可以获得大突变的菌株,因此它也成为发酵工程中的一种有效而安全的育种工具[5]。
原生质体制备和再生是基因组重排技术中非常重要的两个环节[6]。目前关于高产核苷磷酸化酶菌株选育的报道很多,但通过基因组重排技术提高短乳杆菌产胸苷磷酸化酶的报道还未见。本实验系统的对产胸苷磷酸化酶短乳杆菌的原生质体制备及再生条件进行优化,并利用紫外-加热灭活原生质体融合的方法获得短乳杆菌融合子且对融合菌株的性质进行初步分析[7],为进一步通过基因组重排技术选育高产胸苷磷酸化酶短乳杆菌奠定基础。
1 材料与方法
1.1 材料
1.1.1 菌种
亲本:短乳杆菌 E2A(菌体产量高)、短乳杆菌E2B(胸苷磷酸化酶活高),微生物实验室保藏。
1.1.2 培养基及溶液
种子培养基(g/L):酵母膏10,双蒸水1 000 mL,pH 7.0~7.5。
发酵培养基(g/L):酵母膏10,葡萄糖20,NaCl 5,双蒸水1 000 mL,pH 7.0 ~7.5。
普通固体培养基:在发酵培养基的基础上添加琼脂20 g/L,pH 7.0~7.5。
初筛培养基(g/L):酵母膏10,葡萄糖20,NaCl 5,K2HPO4·3H2O2,胸苷 20 mmol,双蒸水 1 000 mL,pH 7.0~7.5。
再生培养基:普通发酵培养基中添加蔗糖0.5 mol/L,MgCl220 mmol/L,CaCl220 mmol/L,明胶 25 g/L,牛血清白蛋白(0.45 um微孔滤膜除菌)5 g/L,琼脂20 g/L。均调 pH 6.5~7.0,以上培养基在115℃高压灭菌15 min。
1.1.3 试剂
渗透压稳定剂LPB(Tris-HCl 10 mmol/L,MgCl2·2H2O 20 mmol/L,蔗糖 0.5 mol/L),pH 6.5,115℃灭菌15 min。
聚乙二醇(PEG 6000)购自国药集团化学试剂有限公司;其他试剂均为国产分析纯。
1.1.4 酶
溶菌酶(20 000 U/mg solid),国药集团化学试剂有限公司,用LPB配制,0.22 μm微孔滤膜过滤除菌;变溶菌素(11 029 U/mg solid),sigma公司,按照5000 U/mL溶解在0.1 mol/L的磷酸钾缓冲溶液中,pH 6.2,0.22 μm微孔过滤除菌,-20℃保存备用。
1.2 实验方法
1.2.1 菌体培养与收集
从平板上挑取单菌落接种到种子发酵液中,37℃,110 r/min活化10~12 h,同样发酵条件下,按照2%的接种量接入含有0.6%甘氨酸预处理的发酵培养基中,培养至对数生长期,离心去上清液收集湿菌体备用。
1.2.2 原生质体的制备及再生
用含不同浓度甘氨酸的发酵液将短乳杆菌培养至对数生长期,5 000 r/min离心15 min,弃上清液,然后用无菌生理盐水和渗透压稳定剂LPB各洗涤1次,离心去除上清液,收集湿菌体。取菌悬液适当稀释后用血球计数板计数测定菌浓,湿菌体用LPB悬浮保证菌浓在1~10×108CFU/mL,悬浮液中加入不同浓度的溶菌酶或变溶菌素[8-9],置于不同温度恒温水浴酶解一定时间,镜检观察原生质体的形成情况。酶解后菌液3 000 r/min离心10 min收集原生质体,用渗透压稳定剂LPB洗涤1次,最后将原生质体悬浮于渗透压稳定剂LPB中。将菌悬液和原生质体液分别用无菌水和渗透压稳定剂梯度稀释后涂布于普通固体培养基和再生培养基,37℃恒温培养24~36 h,测定短乳杆菌原生质体的制备率和再生率[10]。
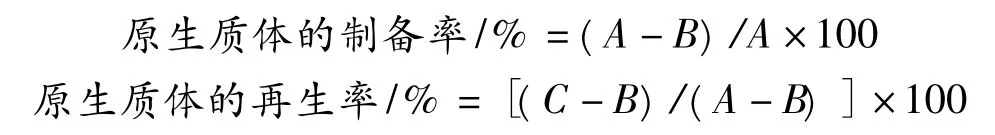
式中:A为酶解前的总菌落数;B为酶解后未脱壁菌体菌落数;C为酶解后原生质体与未脱壁菌体的再生菌落数之和。
1.2.3 短乳杆菌原生质体UV-加热灭活条件的确定
亲本E2A和E2B原生质体液等比例混合,调整菌浓为108CFU/mL。取10 mL亲本原生质体液于培养皿(φ90 mm)中,置于磁力搅拌器上,用20 W紫外灯距离20 cm照射,取不同照射时间的处理液涂布于再生培养基平板上培养,确定紫外致死时间。另取10 mL亲本原生质体液于无菌离心管中,置于60℃水浴中热灭活,取不同加热时间的处理液涂布于再生培养基上培养,确定热灭活时间。
1.2.4 原生质体融合
根据“致死损伤融合互补”原理[11-13],将紫外灭活和热灭活的原生质体等量混合,3 000 r/min离心10 min,去上清液,将原生质体重悬于0.5 mL渗透压稳定剂 LPB中,加入4.5 mL预热至30℃的40%PEG 6000,充分混匀,于30℃水浴中恒温融合10 min,再加入5 mL渗透压稳定剂LPB终止融合,随即离心(3 500 r/min,5 min)洗涤1次,再用LPB适当稀释后,取100 μL涂布于再生培养基上,37℃恒温避光培养48 h筛选融合子。同时分别用紫外灭活和热灭活的原生质体单独融合做对照观察融合情况。
1.2.5 短乳杆菌融合子初筛方法建立
将再生培养基上生长旺盛的融合子菌落分别接种到普通固体培养基和初筛培养基上,初筛培养基在基础发酵培养基的基础上添加底物胸苷和磷酸根,胸苷磷酸化酶为胞内酶,短乳杆菌在生长的过程中,菌体可以利用体内胸苷磷酸化酶催化发酵培养基中的胸苷和磷酸根生成胸腺嘧啶,胸腺嘧啶的生成使得发酵液在290 nm处吸光值增加,从而可粗略判断菌体产胸苷磷酸化酶能力。将初筛培养液发酵10 h后分别用无菌水和pH 13的NaOH稀释10倍和100倍测定其在600 nm和290 nm处吸光值。挑取OD600和OD290较高的融合菌体继续进行胸苷磷酸化酶活测定。
他反反复复地练习滑翔,早早便成为了云浮“飞”得最远的人。但那种借助外物机械带来的飞行体验,又怎能满足得了他的心呢?他无数次地坐在山巅,仰望无尽的苍穹,望着云浮山上空飞过的雄鹰和鸿雁,望着云浮山下飞过的雨燕和黄鹂,他无数次地想,如果能飞一次,哪怕只有一次,此生也便无憾事了。
1.2.6 胸苷磷酸化酶活测定
据Saunders等人报道[14],采用紫外分光光度法测定反应生成的胸腺嘧啶来表征产酶量。标准酶反应混合液包含一定浓度的湿菌体,25 mmol/L的胸苷,1 mmol/L 的 EDTA,pH 7.3、50 mmol/L 的磷酸钾缓冲溶液。55℃条件下反应一段时间,反应结束后,反应液在沸水中煮沸5 min终止反应,离心去除沉淀。上清液用pH 12的NaOH稀释100倍,然后测定290 nm的紫外吸光度OD290nm的增值[15]。胸苷磷酸化酶酶活单位定义为:在上述条件下,1 min内OD290nm变化0.01所需的湿菌体量定义为短乳杆菌的1 个酶活力单位[4]。
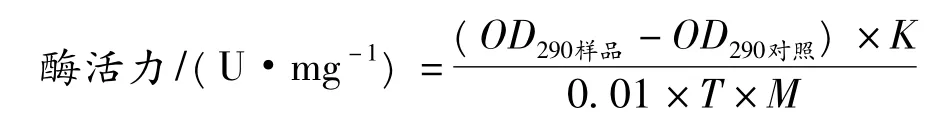
式中:K为稀释倍数(100);T为酶反应时间,min;M为湿菌体质量,mg。
2 结果与分析
2.1 短乳杆菌原生质体制备率和再生率[16]
2.1.1 短乳杆菌生长曲线
为了确定菌体的对数生长期,考察了短乳杆菌在发酵培养基中的生长过程。由图1发现短乳杆菌生长速度较快,培养2 h时开始进入对数生长期,6 h后达到稳定期。不同生长期菌体对溶菌酶的敏感程度有很大差别,处于对数生长后期的菌体细胞壁易被酶解形成原生质体,故选取培养5~6 h的菌体进行原生质体的制备。

图1 短乳杆菌生长曲线Fig.1 Growth curve of Lactobacillus brevis
2.1.2 甘氨酸浓度对短乳杆菌原生质体制备率及再生率的影响
酶解脱壁前添加甘氨酸可以改变细胞结构,使之发生松动增加其对溶菌酶的敏感性,由表1可以发现,短乳杆菌对甘氨酸的敏感性随着浓度的增加而增强,当甘氨酸浓度达到0.9%时,甘氨酸对菌体生长又明显抑制作用,难以获得原生质体,而低浓度的甘氨酸可以明显提高原生质体制备率,对再生率的作用不大,故实验中用含0.6%甘氨酸的发酵液培养菌体至对数生长期。
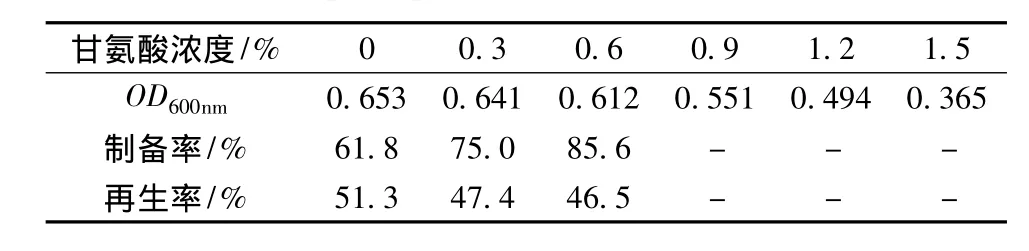
表1 甘氨酸对短乳杆菌原生质体制备率与再生率的影响Table 1 Effects of glycine on formation and regeneration rate of protoplast of Lactobacillus brevis
采用不同质量浓度的溶菌酶和变溶菌素30℃恒温酶解短乳杆菌,其原生质体的制备率和再生率随着溶菌酶浓度的增加,逐渐增加,溶菌酶浓度为2 mg/mL时短乳杆菌原生质体制备率和再生率分别为74.1%和53.6%。当溶菌酶浓度为4 mg/mL时,原生质体制备率高达93.5%,但是再生率仅为22.5%,可能是过高浓度的溶菌酶,水解完细胞壁后,会继续水解细胞膜上的部分蛋白质从而导致细胞死亡,致使原生质体不能再生。在2 mg/mL溶菌酶的基础上添加不同浓度的变溶菌素,结果显示变溶菌素对短乳杆菌作用不明显。因此综合考虑原生质体制备率和再生率,使用2 mg/mL溶菌酶酶解短乳杆菌制备原生质体。
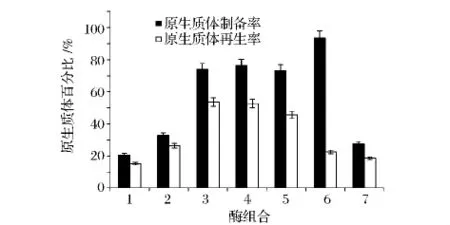
图2 不同酶种类及其质量浓度对短乳杆菌原生质体制备率及再生率的影响Fig.2 Effects of types and concentration of enzymes on formation and regeneration rate of protoplast of Lactobacillus brevis
2.1.4 酶解温度对原生质体制备率和再生率的影响酶解温度可以直接影响到酶促反应速率和原生质体化程度。本实验分别设置30、37、40、45℃ 4个温度梯度,在2 mg/mL溶菌酶酶解90 min条件下进行试验,考察酶解温度对原生质体制备率和再生率的影响,结果见图3。原生质体制备率随温度升高而升高,但原生质体再生率在37℃时出现拐点,达到最大值56.0%。因此本研究选用37℃作为短乳杆菌制备原生质体的最佳酶解温度。
2.1.5 酶解时间对短乳杆菌原生质体制备率和再生率的影响

图3 酶解温度对短乳杆菌原生质体制备率及再生率的影响Fig.3 Effects of enzymolysis temperature on formation and regeneration rate of protoplast of Lactobacillus brevis
将短乳杆菌培养6 h至对数生长后期,2 mg/mL溶菌酶在37℃恒温酶解不同时间,结果如图4。原生质体制备率随着酶解时间的延长而升高,酶解2 h时,原生质体的制备率达到95%,再生率达到48%,若继续延长酶解时间,原生质体制备率可达到100%,但再生率大大降低,故最佳酶解时间为2 h。
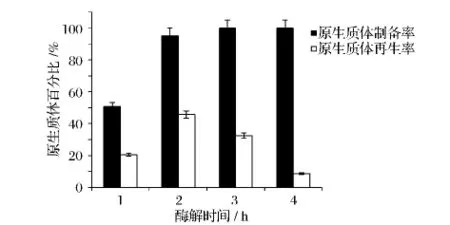
图4 酶解时间对短乳杆菌原生质体制备率及再生率的影响Fig.4 Effects of enzymolysis time on formation andregeneration rate of protoplast of Lactobacillus brevis
2.2 短乳杆菌原生质体UV-加热灭活时间确定
2.2.1 短乳杆菌原生质体的紫外灭活
由表2可知,使用20 W紫外线距离20 cm照射50 min,短乳杆菌原生质体(包括未酶解的菌体)几乎完全灭活,只有约6.2×10-7的低存活率。因此,短乳杆菌原生质体紫外灭活条件为:紫外灯功率20 W,距离20 cm,照射时间为50 min。

表2 短乳杆菌原生质体紫外灭活时间对灭活率的影响Table 2 Effect of UV inactivated time on inactivation rate of Lactobacillus brevis protoplast
2.2.2 短乳杆菌原生质体的热灭活
由于热灭活的原发作用主要在细胞质中,故加热温度不能过高,本研究在60℃条件下对短乳杆菌原生质体加热不同时间以确定其完全致死加热时间。由表2可知,原生质体在60℃水浴中加热60 min,存活率仅为3.2×10-7,故热灭活条件为:60℃水浴加热60 min。

表3 短乳杆菌原生质体热灭活时间对灭活率的影响Table 3 Effect of heating inactivated time on inactivation rate of Lactobacillus brevis protoplast
2.3 融合子的筛选
紫外使细胞致死的原发作用点主要在DNA上,热灭活的原发作用主要在细胞质中,双亲灭活原生质体融合后获得重组体的机制是细胞致死率损伤经过融合得以互补的结果,使用双亲灭活融合获得重组子可以减少对亲株的遗传标记工作,使融合子的检出变得直观,提高筛选融合子的效率[11]。分别以紫外灭活和加热灭活的原生质体单独做融合观察双亲灭活原生质体融合情况。
2.3.1 融合子初筛
前期试验发现,20 mmol/L胸苷和2 g/L磷酸根对短乳杆菌菌体生长影响不显著,短乳杆菌可以直接利用发酵培养基中的胸苷和磷酸根在其胸苷磷酸化酶的作用下生成胸腺嘧啶,从而使发酵液在290 nm出的吸光值增加,图5为融合子初筛结果,短乳杆菌原生质体经过融合后菌体产量和酶活都稍有提高,融合子F15、F16、F17产菌体量和酶活提高较其他融合子显著,故对这3株融合子继续进行酶活测定及菌株稳定性测试。

图5 融合子菌体生长及胸苷磷酸化酶活初筛试验结果Fig.5 Results of cell growth and thymidine phosphorylase for the fusant
2.3.2 融合子F15、F16、F17酶活测定及遗传稳定性测定
初筛挑选出的融合菌株F15、F16、F17分别测试其菌体发酵量和胸苷磷酸化酶活,以亲本E2A和E2B为对照,测试结果如表4。从表4可以看到,融合子菌体发较量和胸苷磷酸化酶活均有提高,其中F15酶活提高50%,达到1.538 U/mg湿菌体,菌体发较量也略有提高,对融合子F15五次传代后,改菌株产胸苷磷酸化酶活稳定在1.500 U/mg湿菌体左右。
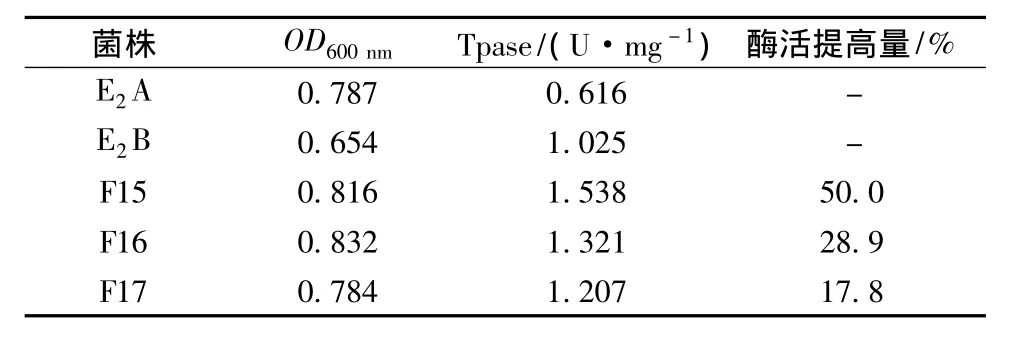
表4 产胸苷磷酸化酶亲本E2A、E2B和融合菌株的酶活性能比较Table 4 Comparison of thymidine phosphorylase for parent strains E2A,E2B and fusants
3 结论
3.1 原生质体的制备与再生
合适的菌龄、菌体浓度及酶解条件是原生质体制备与再生成功的前提条件[17]。对数期的菌体生理状态一致,代谢旺盛,菌体活性高,对酶的敏感性强,故易于原生质体化与再生;不同种属的微生物在制备原生质体时所需的酶种类和浓度不同。本实验发现短乳杆菌对变溶菌素不敏感,对溶菌酶较敏感,2 mg/mL的溶菌酶有利于短乳杆菌原生质体的制备与再生;酶解温度可以直接影响到酶促反应速率和原生质体化程度,实验发现溶菌酶酶解短乳杆菌细胞壁的最适酶解温度为37℃;酶解时间对原生质体的形成和再生起着重要作用,酶解时间太短,细胞壁脱壁不完全,易再生,但原生质体形成率低,酶解时间过长,脱壁完全,原生质膜易被酶液破坏,原生质体再生率低,不利于融合子的形成,综合以上影响因素确定短乳杆菌原生质体制备与再生的较佳条件为:菌体发酵至对数生长中后期,调整菌浓为108CFU/mL时,用2 mg/mL溶菌酶在37℃恒温水浴中酶解2 h,此时原生质体的制备率和再生率分别达到95%和48%左右,有利于后续原生质体融合及基因组重排等技术开展[18]。
3.2 短乳杆菌原生质体融合及融合子性质
与传统诱变相比,原生质体融合具有集中双亲本优良性状的优势,原生质体选育优良性状融合菌株的方法已有很多研究。工业微生物杂交育种必须对亲株进行遗传标记,本研究利用双亲灭活的方法对原生质体进行融合,可以省去亲本遗传标记这步繁琐的工作。文献资料显示,双亲皆用UV灭活仍有一定的重组率,皆用热灭活时,重组率为零。根据“致死损伤融合互补”原理,对亲本原生质体分别进行UV灭活50 min,热灭活60 min,用40%化学融合剂PEG 6000在30℃水浴中融合10 min,挑取再生培养基上的融合子进行初筛和复筛,融合菌株的菌体发酵量和产胸苷磷酸化酶的能力均有提高,融合子F15的酶活达到1.538 U/mg湿菌体,经过5次传代后进行酶活测试,酶活均维持在1.500 U/mg湿菌体,具有良好的遗传稳定性。
[1] 李晓辉,孙嘉康,高彤等.嘌呤核苷磷酸化酶基因的克隆及原核表达载体的构建[J].工业微生物,2006,36(3):27-31.
[2] 邱蔚然,丁庆豹.酶法合成核苷类抗病毒药物[J].中国医药工业杂志,1999,30(10):474-479.
[3] 丁庆豹.核苷磷酸化酶的克隆、表达与应用[D].上海:华东理工大学,2010:56-60.
[4] 谭黎,欧阳立阳,丁庆豹.大肠杆菌核苷磷酸化酶的重组表达和活性[J].华东理工大学学报,2008,24(5):660-664.
[5] 贾建波,陈军,时号.乳酸菌融合菌的构建及其特性研究[J].食品科学,2009,30(21):309-311.
[6] 毛雨,王丹,李强等.产琥珀酸放线杆菌的原生质体制备与再生[J].中国生物工程杂志,2010,30(6):103-108.
[7] 王玉华,李岩,斐晓林,等.基因组改组提高干酪乳杆菌耐酸性生产L-乳酸[J].中国生物工程杂志,2006,26(2):53-58.
[8] Mata S,Dipti J,Shalaka G,etal.Protoplast formation and regeneration inLactobacillus delbruekii[J].Indian J Microbiol,2010,50(1):97 -100.
[9] Lei Y,Xiao P,Ting L,et al.Genome shuffling enhanced l-lactic acid production by improving glucose tolerance ofLactobacillus rhamnosus[J].Journal of Biotechnology,2008,10(1):154-159.
[10] 王慕华,孙文敬,郭金权,等.紫外诱变原生质体选育 D-核糖生产菌株[J].工业微生物,2005,35(1):24-27.
[11] 李焕楼,陈正道.新霉素产生菌弗氏链霉菌灭活原生质体的融合[J].抗生素,1984,9(6):445-449.
[12] Fodor K,Demiri E,Alfoldi L.Polyethylene glycol-induced fusion of heat inactivated and living protoplasts ofBacillus megaterium[J].Bacteriology,1978,135(1):68-70.
[13] Fodor K,Alfoldi L.Polyethylene-glycol induced fusion of bacterial protoplasts[J].Molecular and General Genetics,1979,168(1):55 -59.
[14] Saunders P P,Wilson A B,Saunders G F,et al.Purification and Comparative Properties of a Pyrimidine Nucleoside Phosphorylase fromBacillus stearothermophilus[J].The Journal of Biological Chemistry,1969,244(13):3 691-3 697.
[15] 魏晓琨,张春艳,丁庆豹,等.产气肠杆菌菌体内核苷磷酸化酶酶活测定条件的分析[J].中国临床志,2007,175(12):4 -7.
[16] 周东坡,平文祥.微生物原生质体融合与基因组重排[M].北京:中国科学技术出版社,2010:139-177.
[17] Hopwood D A,Wright H M,Bill M J,et al.Genetic recombination through protoplast fusion inStreptomyces[J].Nature,1979,268(14):171 -174.
[18] Ranjan P,Susan L,Vesna G.Genome shuffling ofLactobacillusfor improved acid tolerance[J].Nature Biotechnology,2002,20(10):702 -707.
