恒电流刺激下神经元Chay模型的Hopf分岔分析
2013-10-26李洪明张素丽
李洪明,张素丽
(1.呼伦贝尔学院 数学系,内蒙古 呼伦贝尔 021008;2.太原理工大学 数学学院,太原 030024)
1 Chay模型的概述
20世纪50年代Hodgkin和Huxley首次提出了经典Hodgkin-Huxley模型[1](简称 H-H模型),该方程主要研究神经元细胞的放电行为。但是HodGkin和Huxley主要考虑的是神经轴突膜中的K+和Na+通道离子电流,因而不能全面描述神经元细胞的膜电压特性。1994年,Chay等人通过大量实验,在H-H模型的基础上提出Chay模型[2]。Chay模型除了考虑可兴奋细胞膜上Na+和K+通道及漏电流通道的作用外,还有赖于Ca2+浓度的K+离子通道,后者对于依赖Ca2+的各类肌细胞和胰岛B细胞的放电是极重要的。因此神经元Chay模型能更好地模拟神经元的各种放电形式。
古华光等主要研究了Chay模型的两类跃迁机制产生的整数倍神经放电节律[3-7];崔睿等研究了三态跃进机制产生的两种放电节律[8];张春燕等进行了神经元典型放电的计算机仿真[9];裴利军等进行了神经元Chay模型的动力学分析[10];陆启韶等对神经元Chay模型的生理特性进行了深入的详细研究[12-14]。本文主要利用非线性动力学方法对Chay模型进行了单参数的Hopf分岔分析,然后证明了其Hopf分岔的存在性。
Chay模型可用下面的三阶非线性微分方程表示[2]:
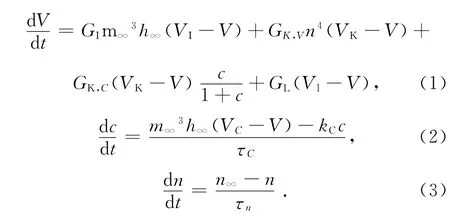
方程(1)描述了神经元细胞的膜电位的变化规律;VI,VK,VL,V 分别表示 Na+-Ca2+通道、K+通道、漏电流的电位和平衡电位;GI,GK,V,GK,C,GL分别代表各通道的最大电导;c为细胞内Ca2+浓度;n为K+通道开放概率;m∞,h∞分别是Ca2+、Na+通道门打开的概率。
方程(2)描述了细胞膜内Ca2+变化规律。VC是Ca2+可逆电位;τc为常量,τc=100/27;kC表示Ca2+流出细胞内的速率常数。
方程(3)描述了K+通道门打开概率的变化规律。τn是弛豫时间常数;n∞为K+通道门打开的概率。
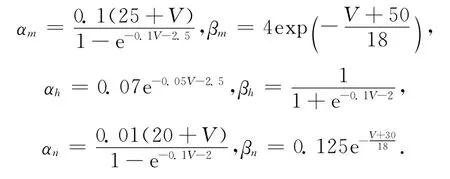
由文献[2]知,m∞,n∞,h∞分别是Ca2+、K+和Na+离子通道门打开的概率,为
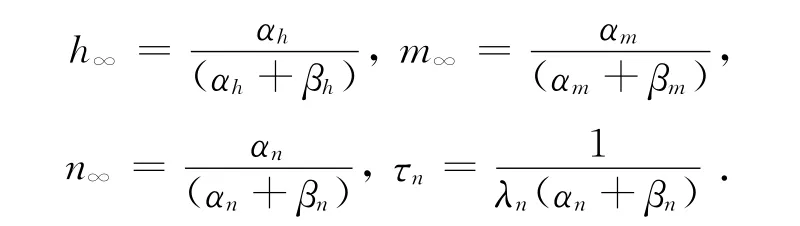
本文中参数取值如下:GI=1800S;GK,V=1700S;GL=7S;VI=100mV ;VK=-75mV;VL=-40mV;V=100mV
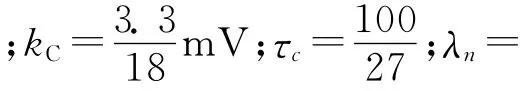
2 Chay模型的平衡点及其稳定性
对于方程(1)、(2)、(3)所示的自治系统,设其平衡点为A(Ve,ce,ne),令方程(1)(2)(3)右端等于0,即
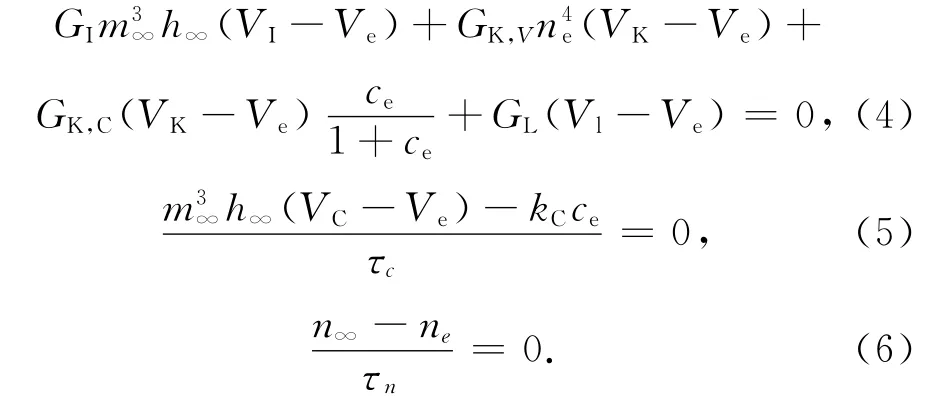
由式(5)、(6)可知:
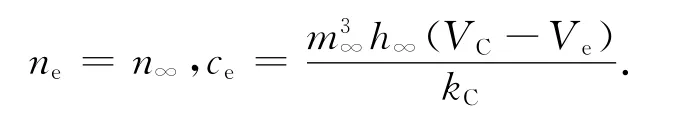
则ne,ce均为Ve的函数。将ne,ce代入(4)得:
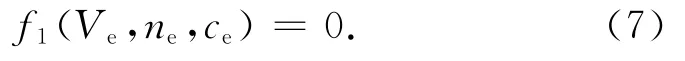
数值计算发现,式(7)仅有一个解,得平衡点为

Chay模型在该点处的Jacobian矩阵为

其相应的特征根为

根据稳定性理论[11],由于平衡点的特征根有正实部,可以判定平衡点A(Ve,ce,ne)为不稳定的点。
3 恒电流刺激下神经元Chay模型的Hopf分岔分析
恒电流刺激下Chay模型可用下面的三阶非线性微分方程表示:
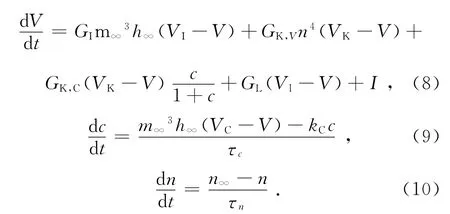
式中,I为恒流电流。
设恒电流刺激下系统的平衡点为B(Ve,ce,ne),模型参数取已给出的值,改变I的值,通过数值计算方法求解,得到如表2所示。

表1 其他参数不变,只改变I时,Chay模型的唯一平衡点 B(Ve,ce,ne)的坐标、特征根及其稳定性
恒电流刺激下神经元Chay模型在平衡点B(Ve,ce,ne)处的Jacobian矩阵为J(B),数值计算得到系统在I取不同值时的特征值,如表1所示。前三种情况,即I分别为-65,-66,-66.6μA/cm2时,均存在正实部特征根,根据稳定性理论[11],可以判定平衡点B(Ve,ce,ne)为不稳定的结点。后2种情况,即I分别为-66.7,-66.8,-67μA/cm2时,其特征根为一对共轭复根与一实根,且共轭复根的实部与实根都为负,则可判定其为渐进稳定的结点。
4 数值模拟
以上的理论计算得到的分岔情况只反映了参数变化引起的系统稳态解的变化。为了全面考察参数对模型的动态特性的影响,下面用Matlab软件进行了参数连续变化下的数值模拟。图1、2、3验证了当I=-65,I=-66,I=-66.6μA/cm2时,神经纤维产生周期性动作电位,Chay模型出现稳定周期解。
图4、5显示了当I通过-66.6μA/cm2时,在其附近发生Hopf分岔,其跨膜电压经小幅衰减振荡后稳定在略高于稳定点的某一平衡点,周期解消失,可以看到,随着I的增大,当越过临界值时,系统出现渐进稳定平衡点,振荡消失。
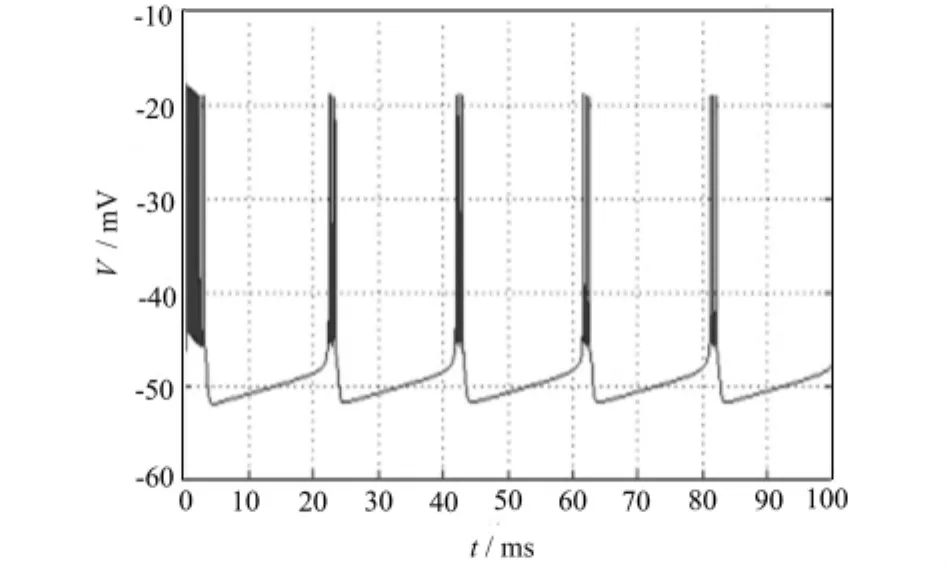
图1 I=65μA/cm2时膜电位V随时间t的变化
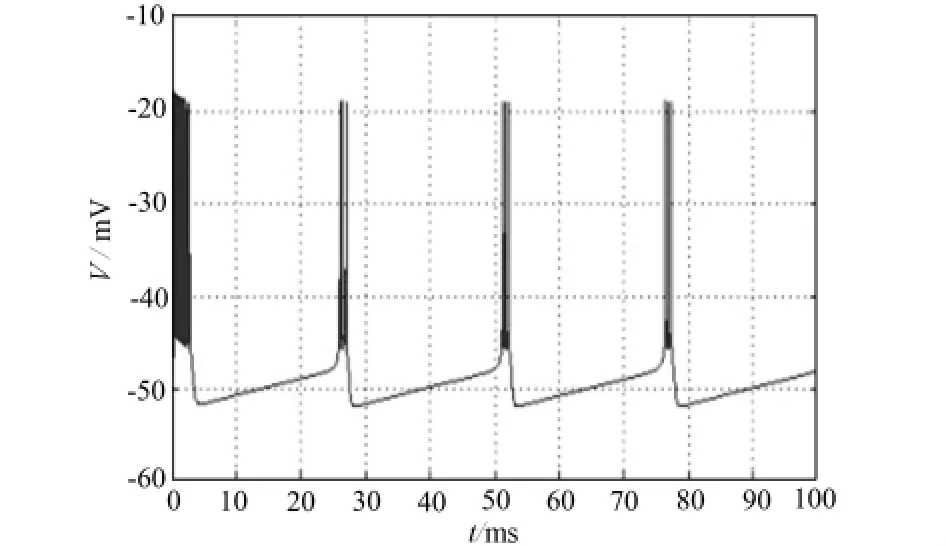
图2 I=-66μA/cm2时膜电位V随时间t的变化
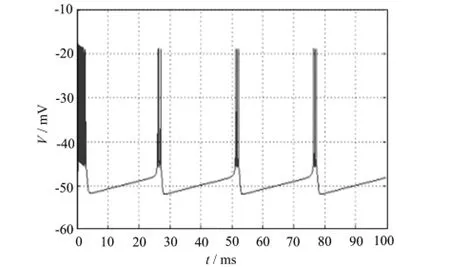
图3 I=-66.6μA/cm2时膜电位V随时间t的变化
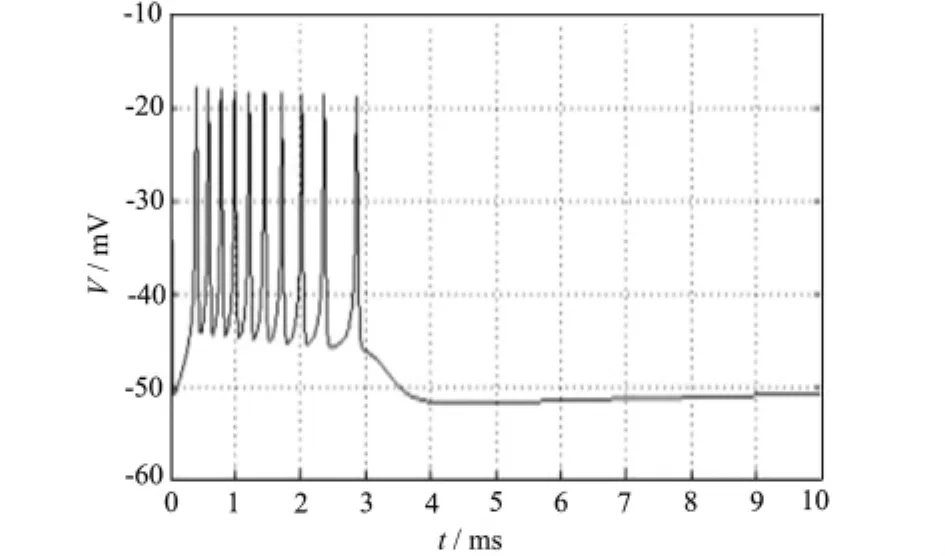
图4 I=-66.7μA/cm2时膜电位V随时间t的变化

图5 I=-67μA/cm2时膜电位V随时间t的变化
5 结束语
本文仅对恒电流刺激下神经元Chay模型的Hopf分岔进行了分析,与实际生理系统中多因素的共同作用有较大差别,因此还需进一步的对全局分岔和多层次分岔结构进行研究。此外,实际神经系统中的神经元总是受到内外噪声、参数的微小涨落等影响。因此,我们需考虑更多的随机因素的作用。
[1]Hodgkin A L,Huxley A F.A quantitative description of membrane current and its application to conduction and excitation in nerve[J].J Physiol,1952,117:500-544.
[2]Chay T R.Chaos in three variable model of an excitable cell[J].Physica D,1985,16(2):233-242.
[3]杨明浩,古华光,李莉,等.神经放电加周期分岔中由随机自共振引起的一类新节律[J].生物物理学报,2004,20(6):465-470.
[4]古华光,李莉,杨明浩,等.实验性神经起步点产生的整数倍簇放电节律[J].生物物理学报,2003,20(1):68-72.
[5]古华光,任维,杨明浩,等.神经自发整数倍峰放电节律的随机性和确定性模式的比较[J].生物物理学报,2003,19(3):272-278.
[6]古华光,任维,陆启韶,等.实验性神经起步点自发放电的分叉和整数倍节律[J].生物物理学报,2001,17(4):637-644.
[7]Gu HG,Ren W,Lu QS,et al.Integer multiple spiking in neuronal pacemakers without external periodic stimulation[J].Phys Lett A,2001,285:63-68.
[8]崔睿,王珊珊,李晋,等.由三态跃进机制产生的两种神经放电节律[J].生物物理学报,2011,27(8):703-711.
[9]张春燕,田心.可兴奋神经元典型放电的计算机仿真[J].天津医科大学学报,2004,(s1)
[10]裴利军,王永刚,范烨.神经元Chay模型的动力学分析[J].郑州大学学报(理学版),2009,2(41):7-12.
[11]马知恩,周义仓.常微分方程定性与稳定性方法[M].北京:科学出版社,2001:41-92.
[12]Duan LX,Lu QS.Codimension-two bifurcation analysis on firing activities in Chay neuron model[J].Chaos,Solitions and Fracials,2006,30(5):1172-1179.
[13]Yang ZQ,Lu QS.Transitions from bursting to spiking due to depolarizing current in the Chay neuronal model[J].Communications in Nonlinear Science and Numerical Simulation,2007,12(3):357-365.
[14]Duan LX,Lu QS,Wang QY.Two-parameter bifurcation analysis of firing activities in the Chay neuronal model[J].Neurocomputer,2008,72(1-3):341-351
