基于线粒体基因序列和齿舌对古氏非螺分类地位的探讨
2013-10-13李海涛时小军周俊杰吕向立
李海涛, 时小军, 周俊杰, 周 鹏, 吕向立
(国家海洋局 南海环境监测中心, 广东 广州 510300)
几个世纪以来, 贝壳形态一直是腹足类动物最主要的分类依据[1]。传统的分类学者主要是基于贝壳最主要的几个特征来对物种进行分类。但这些特征在进行科、属等较高阶元划分时易受到分类者主观认识的影响[2]。
分子生物学技术的迅速发展, 为解决经典分类中的疑难问题提供了有力的帮助。线粒体DNA因其分子结构简单、严格母系遗传、几乎不发生重组、进化速率快等优点, 广泛应用于海洋腹足类分子系统发育研究中[3-6]。
古氏非螺(Afer cumingii(Reeve, 1844))是一种在我国较为少见的海洋腹足类, 以前仅在台湾地区有过报道, 大陆一直未采到标本。因未做过系统研究,该种在中国大陆和日本被归为犬齿螺科(Vasidae)[7-10], 中国台湾[11]将本种归入拳螺科(Turbinellidae)。而根据齿舌的构造, Fraussen 等[12]将非螺属(Afer)修订到蛾螺科(Buccinidae)内, 但这一结论至今仍未被广泛采用。以往的形态分类, 可能是因非螺属贝壳轴唇上具有褶襞这一特征, 而被归为犬齿螺科或拳螺科[12]。到目前为止, 非螺属的分类地位还缺乏分子证据的支持。本研究基于线粒体 CO1和16S rRNA基因序列, 并依据齿舌形态, 探讨了古氏非螺的分类地位, 为进一步证实非螺属的分类地位提供分子证据。
1 材料与方法
1.1 实验材料
本研究所用的古氏非螺标本共 5个, 其中两个为活体, 标本采自南海北部陆架区, 水深140~176 m。活体标本于-20℃冷冻保存带回实验室后解剖其齿舌,肌肉组织保存于90%乙醇溶液中备用。
1.2 DNA提取、PCR扩增
取样品腹足肌100 mg左右, 采用CTAB法提取基因组DNA, 酚/氯仿纯化, -20℃保存备用。
扩增 CO1基因的引物为: LCO-1490 5’- GGT CAA CAA ATC ATA AAG ATA TTG G-3’和 LCO-2198 5’-TAA ACT TCA GGG TGA CCA AAA AAT CA-3’[13];扩增 16S rRNA基因的引物为 16sar-L 5’-CGC CTG TTT ATC AAA AAC AT-3’和 16sbr-H 5’-CCG GTC TGA ACT CAG ATC ACG T-3’[14]。
PCR 反应体系为 50 μL, 包括 10×buffer(含Mg2+)5 μL, dNTP 200 μmol/L, 引物各 0.2 μmol/L,Taq酶1.25 U, DNA模板1 μL, 灭菌蒸馏水补足至50 μL。PCR反应条件为: 94℃预变性3 min, 然后进行35个循环, 每循环包括94℃1 min, 50℃1 min, 72℃1 min, 最后72℃延伸10 min。PCR产物经电泳检测后,送上海生工生物工程有限公司进行纯化并双向测序。
1.3 序列分析
利用DAMBE软件对GenBank中部分已知蛾螺科种类的序列与本研究中测定的序列进行多序列比对。以犬齿螺科种类为外群, 采用MEGA 4.1软件以邻接法(neighbor joining, NJ)和最大简约法(maximum parsimony, MP)构建分子系统发育树, 系统树各分支的置信度由Bootstrap l000循环检验。大于50%的Bootstrap支持率标注在系统树上。
2 结果
2.1 贝壳及齿舌特征
贝壳呈长纺锤形, 螺层约7层; 壳高65~76 mm,壳宽27~30 mm; 壳表黄褐色, 具有深褐色斑点和斑块, 并雕刻有明显的细螺肋; 体螺层膨大; 每一螺层的中部扩张形成肩角, 肩角上有一列发达的结节状突起, 体螺层的肩角上具有10~12个; 壳口大, 卵圆形, 内淡褐色或白色; 前水管沟长, 半管状, 末端常弯曲; 轴唇上有一明显的褶襞; 厣角质, 黄色, 周缘光滑(图 1)。

图1 古氏非螺的贝壳Fig. 1 Shells of Afer cumingii
齿舌为典型的蛾螺科类型, 齿式 0·1·1·1·0, 无缘齿。侧齿笔架状, 具3枚发达的齿尖, 曲向内侧, 外侧一枚最为发达, 形似镰刀; 中央齿基板元宝形, 在中部弯曲并加厚, 其上具3枚齿尖, 呈“山”字形, 中间一枚齿尖最发达(图2)。
2.2 序列特征
测序结果提交 GenBank获得登录号: 两条 16S rRNA基因序列为 JX469112和 JX469113; 两条CO1基因序列为JX469114和JX469115。两个古氏非螺个体CO1和16S rRNA 基因片段的长度分别为658 bp 和509 bp(不包括引物区), 无碱基的插入/缺失。CO1基因片段A, T, G, C的比例分别为26.3%,37.2%, 19.1%和17.3%; 第三位密码子A, T, G, C的比例分别为41.8%, 42.7%, 8.6%和6.8%, A+T的比例显著高于G+C的比例。两条CO1基因序列存在2个(0.3%)碱基的差异, 均为密码子第三位碱基, 未造成氨基酸序列的改变。16S rRNA基因片段A, T, G, C的比例分别为34.4%, 30.5%, 19.6%和15.5%; 两条序列亦存在2个(0.4%)碱基的差异。CO1和16S rRNA基因片段的A+T的比例均明显高于G+C的比例。

图2 古氏非螺的齿舌Fig. 2 Radular teeth of Afer cumingii
2.3 系统学分析结果
本文基于CO1和16S rRNA 基因序列分别构建了 NJ和 MP树, 但在文中仅给出 NJ树, NJ树的Bootstrap支持率标注在节点斜线左侧, MP树的Bootstrap支持率标注在节点斜线右侧。基于CO1基因构建的 NJ和 MP树的拓扑结构完全一致, 基于16S rRNA基因构建的两种树的拓扑结构存在一定的差异。基于两个基因构建的系统发育树都显示, 古氏非螺都与蛾螺科种类聚类, 且支持率较高。在基于CO1基因序列构建的系统树中, 古氏非螺处于较为基部的位置(图3)。在基于16S rRNA基因序列构建的系统树中, 古氏非螺与Buccinulum linea聚为一支, Bootstrap支持率在80%以上(图4)。
3 讨论
齿舌是软体动物重要的内部器官, 也是形态学分类的重要依据之一。如依据齿舌的形态和结构,Quadrasia属从平轴螺科(Planaxidae)修订到蛾螺科[15],Benthovoluta属从涡螺科(Volutidae)修订到拳螺科(Turbinellidae)[16]。古氏非螺的齿舌中央齿和侧齿均具有 3枚齿尖, 符合张玺[17]和蔡英亚等[18]对蛾螺科齿舌的描述。蛾螺科与织纹螺科和盔螺科在亲缘关系上较近, 三科种类的齿舌形态也最为相似。依据中央齿的形态和齿尖数目, 织纹螺科较容易与其他两科的种类区分开来。但根据张玺[17]对蛾螺科和盔螺科的齿舌的描述, 两者的差异并不十分明显, 盔螺科“中央齿几乎永远具3个齿尖, 侧齿通常具两个不同的齿尖”, 一些蛾螺科种类, 如Antillophos属种类的齿舌就符合这一特点[19]。因此, 仅依据齿舌的形态有时候并不能准确判断一些种类的分类地位, 还需要其他证据的支持。
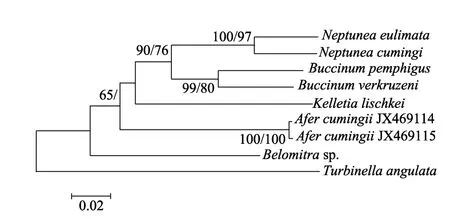
图3 基于CO1基因序列通过邻接法构建的系统发育树Fig. 3 Molecular phylogenetic tree based on CO1 gene using NJ method
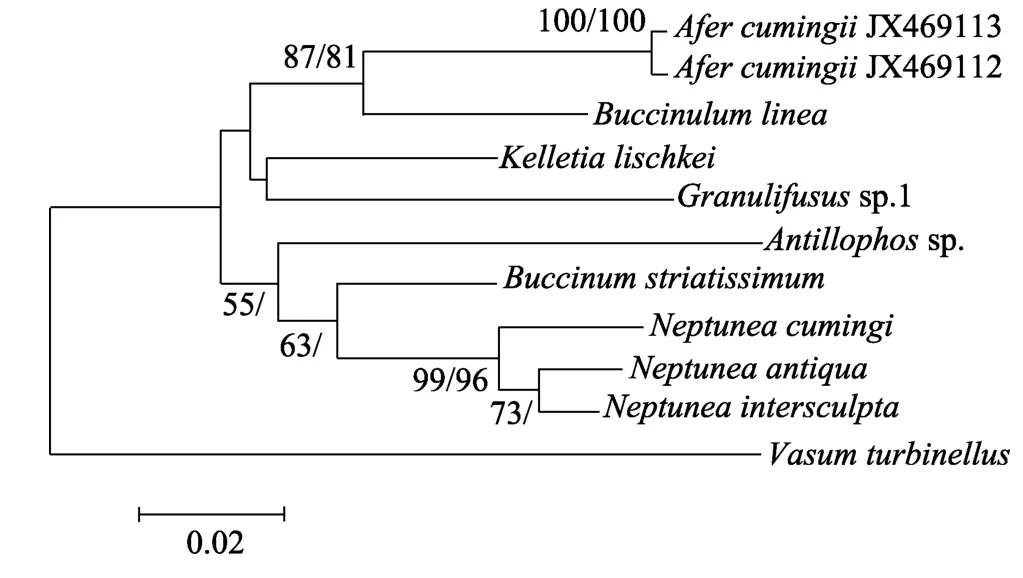
图4 基于16S rRNA基因序列通过邻接法构建的系统发育树Fig. 4 Molecular phylogenetic tree based on 16S rRNA gene using NJ method
本文首次从分子水平证实了古氏非螺或非螺属的分类地位。基于CO1和16S rRNA两个基因片段构建的系统发育树都显示古氏非螺属于蛾螺科, 支持Fraussen等[12]的观点。造成16S rRNA基因片段NJ和 MP树拓扑结构不一致的原因, 可能是该基因片段信息量较少的缘故。董长永等[20]基于核 28S rRNA基因分析了部分蛾螺科种类的系统关系, 结果表明香螺属(Neptunea)是蛾螺科中较为进化的种类,这与本文的结果一致。CO1基因的进化速率比 16S rRNA更快[21], 具有更多的系统发生信息, 但由于本文研究的个体较少并未体现这一点。董长永等[20]基于 28S rRNA基因的变异程度认为水泡蛾螺(Buccinium pemphigum)和黄海蛾螺(B. yokomaruae)可能为同一个种或亚种。但28S rRNA基因高度保守,不适合种类的区分鉴定[22], 这种推断可能并不准确。田莹等[23]的研究结果表明, 水泡蛾螺和黄海蛾螺的齿舌具有明显的差异。将分子标记和形态鉴定相结合, 建立物种间的系统发育关系, 是解决物种分类和鉴定难题的必然途径[24]。
[1]Puillandre N, Cruaud C, Kantor Y I. Cryptic species inGemmuloborsonia(Gastropoda: Conoidea)[J]. Journal of Molluscan Studies, 2010, 76: 11-23.
[2]张爱菊, 尤仲杰. 分子生物技术在在贝类鉴定和分类上的应用[J]. 宁波大学学报(理工版), 2005, 18(3):404-409.
[3]Hayashi S. The molecular phylogeny of the Buccinidae(Caenogastropoda: Neogastropoda)as inferred from the complete mitochondrial 16S rRNA gene sequences of selected representatives [J]. Molluscan Research, 2005,25(2): 85-98.
[4]王莹, 苏成勇, 潘鸿春, 等. 基于线粒体CO1基因序列分析宝贝科主要类群的系统发生关系[J]. 动物分类学报, 2007, 32(1): 124-130.
[5]张爱菊, 尤仲杰. 基于 16SrDNA序列片段探讨织纹螺的系统发生关系[J]. 动物分类学报, 2008, 33(3):549-552.
[6]Modica M V, Bouchet P, Cruaud C, et al. Molecular phylogeny of the nutmeg shells (Neogastropoda,Cancellariidae)[J]. Molecular Phylogenetics and Evolution, 2011, 59(3): 685-697.
[7]李荣冠, 江锦祥. 前鳃亚纲[M]//黄宗国. 中国海洋生物种类与分布. 北京: 海洋出版社, 2008: 490.
[8]张素萍, 李宝泉. 软体动物门[M]//刘瑞玉. 中国海洋生物名录. 北京: 科学出版社, 2008: 500.
[9]李荣冠, 黄宗国. 腹足纲[M]//黄宗国, 林茂. 中国海洋生物图集(第四册). 北京: 海洋出版社, 2012: 120.
[10]菱田嘉一. 世界海产贝类大图鉴[M]. 东京: 久美株式会社, 2000: 250.
[11]赖景阳. 台湾自然观察图鉴(13): 贝类[M]. 中国台湾: 度假出版社, 2000: 104.
[12]Fraussen K, Hadorn R. Transfer ofAferConrad, 1858 to Buccinidae (Neogastropoda)with description of a new species from Western Africa [J]. Gloria Maris,2000, 38 (2-3): 28-42.
[13]Folmer O, Black M, Hoeh W, et al. DNA primers for amplification of mitochondrial cytochrome c oxidase subunitⅠ from diverse metazoan invertebrates [J]. Molecular Marine Biology and Biotechnology, 1994, 3: 294-299.
[14]Simon C, Frati F, Beckenbach A, et al. Evolution,weighting and phlogenetic utility of mitochondrial gene sequences and a compilation of conserved polymerase chain reaction primers[J]. Annals Entomological Society of America, 1994, 87: 651-701.
[15]Houbrick R S. Transfer ofQuadrasiafrom the Planaxidae to the Buccinidae (Mollusca: Gastropoda: Prosobranchia)[J]. Proceedings of the Biological Society of Washington, 1986, 99(2): 359-362.
[16]黑田德米. ツ ノキフデの分 類 学上の位置[J]. 貝類学 雑 誌, 1965, 24(1): 50-52.
[17]张玺, 齐钟彦. 中国经济动物志: 海产软体动物[M].北京: 科学出版社, 1962: 55-60.
[18]蔡英亚, 谢绍河. 广东的海贝(修订版)[M]. 广东:汕头大学出版社, 2006: 65-66.
[19]李海涛, 朱艾嘉, 方宏达, 等. 蛾螺科、织纹螺科和细带螺科腹足类齿舌的形态学研究[J]. 海洋与湖沼,2010, 41(4): 495-499.
[20]董长永, 侯林, 隋娜, 等. 中国沿海蛾螺科5属10种28S rRNA基因的系统学分析[J]. 动物学报, 2008,54(5): 814-821.
[21]苏天凤, 黄建华, 吴进锋, 等. 2种东风螺线粒体基因序列多态性研究[J]. 中国水产科学, 2007, 14(3): 369-376.
[22]喻达辉, 朱嘉濠. 珠母贝属6个种的ITS 2分子标记研究[J]. 南方水产, 2005, 1(4): 6-12.
[23]田莹, 张素萍, 常亚青.黄渤海区蛾螺的齿舌研究[J].海洋科学, 2009, 33(10): 54-58.
[24]张亮, 黄艳艳, 刘焕章. 利用mtDNA 16S rRNA 序列差异鉴定江西青岚湖的河蚌物种[J]. 水生生物学报,2004, 28(3): 294-299.
