CMS-FA 胞质杂交稻不育细胞质和核质互作的遗传效应分析
2013-09-22仇秀丽王洪飞王乃元梁康迳
仇秀丽,王洪飞,王乃元,李 毓,梁康迳
(1.福建农林大学作物遗传育种与综合利用教育部重点实验室,福建福州350002;2.福建省农业科学院水稻研究所,福建福州350018)
水稻细胞质雄性不育材料的利用为杂交稻大面积推广应用奠定了坚实基础.细胞质不仅可以引起水稻的雄性不育,而且还关系到杂交稻产量相关性状的优劣和杂种优势利用的前景[1].因此,对水稻不育细胞质遗传效应的研究受到普遍关注.自1973年我国实现水稻三系配套以来,已育成许多种细胞质来源的水稻雄性不育系,但目前在生产上被广泛应用的不育系细胞质仅为少数几种,杂交籼稻中G型、D型和印尼水田谷型虽有一定的比重,但仍以野败型胞质为主,而杂交粳稻以BT型较多[2,3],不育胞质类型单一、遗传基础狭窄是我国杂交稻生产面临的严峻问题.以扩大杂交稻遗传基础为目的,创造和挖掘可恢复性稳定且无不良细胞质效应的新细胞质源雄性不育遗传资源己成为水稻雄性不育育种的重要方向之一.
福建农林大学新质源杂交稻育种技术研究课题组历经20多年的尝试和探索,成功研究出一种胞质来源于普通野生稻的雄性不育的新材料,定名为CMS-FA[4].研究表明该胞质类型的细胞质育性基因和细胞核育性基因与野败型(CMS-WA)和红莲型(CMS-HL)相互不等位,其恢保关系与CMS-WA和CMS-HL型也不同,是一种全新的雄性不育细胞质类型[4-8].在此基础上育成系列不育系、保持系和恢复系,实现了水稻CMS-FA三系配套.王洪飞等[9]研究表明,CMS-FA胞质杂交稻亲本配合力好,杂种优势明显.为进一步研究CMS-FA胞质杂交稻不育细胞质对其杂种F1产量等相关性状的影响,本研究选用4个CMS-FA胞质不育系、相应保持系和3个恢复系为材料,分析CMS-FA胞质杂交稻不育细胞质对杂种F1产量相关性状的细胞质效应和核质互作效应,为开发利用该胞质类型的杂交稻提供依据.
1 材料与方法
1.1 试验材料
选用CMS-FA胞质不育系金农1A、金农2A、金农3A和新源7A及其相应的保持系为母本,CMS-FA胞质恢复系金恢1号、金恢2号和金恢4号为父本,按NCⅡ杂交方式配制了12对不育胞质杂种(AF1)和可育胞质杂种(BF1).试验材料均由福建农林大学新质源杂交稻育种技术研究课题组提供.
1.2 试验方法
将供试亲本和所有F1于2008年6月5日播种于福建农林大学教学科研基地(沙县)试验田,秧龄均为27 d;试验采用随机区组排列,3次重复.单本栽插,株行距18 cm×18 cm.各小区均种植33株,每区3行,每行11株.试验地前茬水稻,冬季和春季空闲,肥力中等偏上,地力较均匀,施肥和管理参照当地中上水平实施.水稻成熟后,从各小区取5株,考察其株高、单株穗数、穗长、剑叶长、穗颈长、每穗总粒数、每穗实粒数、结实率、千粒重和单株产量等性状.
1.3 统计分析
试验以不育胞质杂种与可育胞质杂种的相对差值(AF1-BF1)/BF1×100%表示不育细胞质的相对效应[10],并进行差异显著性测验.
采用数量性状核基因加性、显性与细胞质互作混合线性模型及其统计分析方法[11-16],检测和分析CMSFA胞质杂交稻核质互作效应及其它们对杂种产量相关性状的影响.用MINQUE(1)法估算CMS-FA胞质杂交稻亲本和杂种F1产量相关性状的核基因加性方差、显性方差、加性或显性与细胞质互作方差分量,并计算各遗传方差分量占表现型方差的比率.利用调整无偏预测法(AUP法)估测CMS-FA胞质杂交稻亲本和杂种F1的核基因加性效应值、显性效应值、加性或显性与细胞质互作效应值.利用群体平均数和各项遗传效应值(核基因加性、显性效应和核质互作效应)预测有关杂种F1产量等相关性状的平均基因型值以及群体杂种优势值.采用Jackknife抽样方法,估算上述各项遗传参数的标准误,用t测验(自由度=2)进行显著性检验.
2 结果与分析
2.1 CMS-FA胞质杂交稻不育细胞质对产量相关性状的影响
两种不同胞质类型杂种的产量相关性状有一定的差异,除株高、穗长和穗颈长3性状外,其他性状是不育胞质杂种的表型值大于可育胞质杂种(表1).不育胞质杂种和可育胞质杂种的平均结实率都大于80%,表明CMS-FA胞质不育系具有良好的可恢复性,展示了CMS-FA胞质杂交稻杂种优势利用的前景.
2.2 CMS-FA胞质杂交稻不育细胞质对产量相关性状的遗传效应
利用细胞质优势率判别细胞质效应,细胞质优势率/%=(AF1-BF1)/BF1×100[10].根据图1可知,不育细胞质对千粒重和生育期表现出显著或极显著的正效应,对穗颈长表现出极显著的负效应,说明细胞质对千粒重、生育期和穗颈长有显著影响.细胞质效应对穗颈长的影响最大(-5.97%),其次是千粒重(1.33%)和生育期(0.52%),CMS-FA细胞质使水稻植株穗颈长显著缩短,千粒重显著提高,生育期显著延长.虽然CMSFA细胞质对单株产量、单株穗数、每穗总粒数、每穗实粒数、结实率、剑叶长等6个性状表现出正效应,对株高和穗长表现出负效应,但相对效应值不显著,表明CMS-F A细胞质对这8个性状影响不明显.
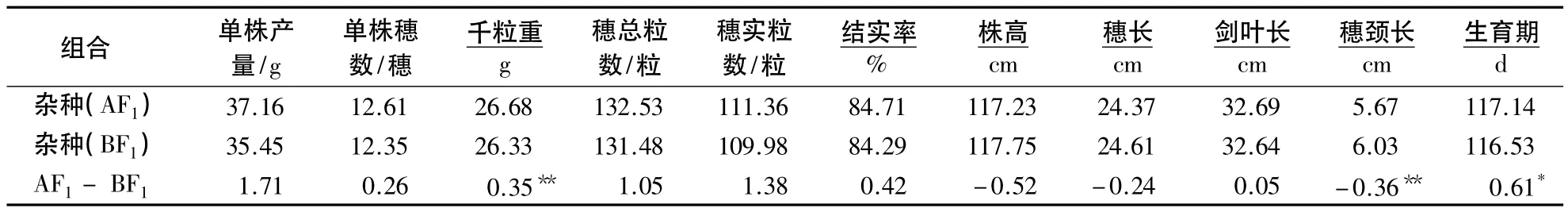
表1 不育胞质杂种(AF1)和可育胞质杂种(BF1)产量相关性状的平均表现1)Table 1 Phenotype means of yield-related traits in two cross types

图1 不育胞质杂种与可育胞质杂种在产量相关性状上的相对效应Fig.1 Relative effect of yield-related traits between sterile cytoplasm and maintainer cytoplasm
2.3 CMS-FA胞质杂交稻产量相关性状的核质互作效应
CMS-FA胞质杂交稻遗传群体中,单株产量和穗颈长的加性方差比率为0,单株穗数的显性方差比率不显著,其他性状的加性方差比率与显性方差比率均达显著或极显著水平,并且在多数性状上加性×细胞质或显性×细胞质互作效应方差比率亦达显著或极显著水平,表明该类遗传群体的各性状核基因×细胞质互作效应是存在的(表2).除穗颈长外,其他性状的VAC/VP和VDC/VP明显小于VA/VP和VD/VP,因此,在CMS-FA胞质杂交稻配组中,上述性状虽然存在明显的核质互作效应,但仍以加性效应和显性效应控制为主.
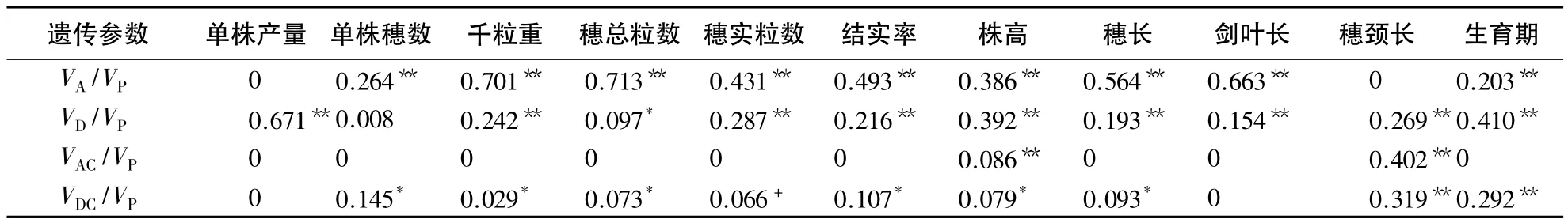
表2 CMS-FA杂交稻产量相关性状遗传方差估算值1)Table 2 Genetic variance estimates for yield-related traits CMS-FA hybrid rice
根据遗传主效应的方差比率分析,单株穗数、千粒重、每穗总粒数、毎穗实粒数、结实率、穗长和剑叶长等性状主要受基因的加性效应控制,而单株产量、株高和生育期则以显性效应控制为主,这与王洪飞等的研究结果基本一致[9].通过比较核基因×细胞质互作效应的方差比率,株高和穗颈长均存在显著或极显著的加性×细胞质与显性×细胞质互作效应,而单株穗数、千粒重、每穗总粒数、结实率、穗长和生育期只存在显著或极显著的显性×细胞质互作效应,说明CMS-FA胞质杂交稻及其后代在这些性状上的表现,除受父母本核基因的作用外,还较为明显地受其核质互作效应的影响.其中,穗颈长受影响的程度最大,其次为生育期和株高,然后依次为单株穗数、结实率、穗长、每穗总粒数和千粒重.试验还表明单株产量、每穗实粒数和剑叶长等性状的核质互作效应不明显.
2.4 不育细胞质的核质互作对CMS-FA胞质杂交稻F1产量相关性状的平均基因型值的影响
通过对上述各项遗传效应值的预测,并以此估算CMS-FA不育胞质杂种(AF1)和可育胞质杂种(BF1)的平均基因型值,根据核基因效应(加性和显性)与核质互作效应进行分解(表3).结果表明,由于不育胞质和可育胞质与核基因互作效应(GC)的影响,使两类水稻新质源杂种的平均基因型值在多数产量相关性状上存在一些差异.核质互作表现为不育系胞质×核基因的互作效应导致杂种F1的千粒重增加和单株产量降低,但程度相对较小,但导致植株增高和穗颈长增长明显.可育胞质×核基因的互作效应导致CMS-FA胞质杂种F1穗长显著增长和穗颈长显著缩短.
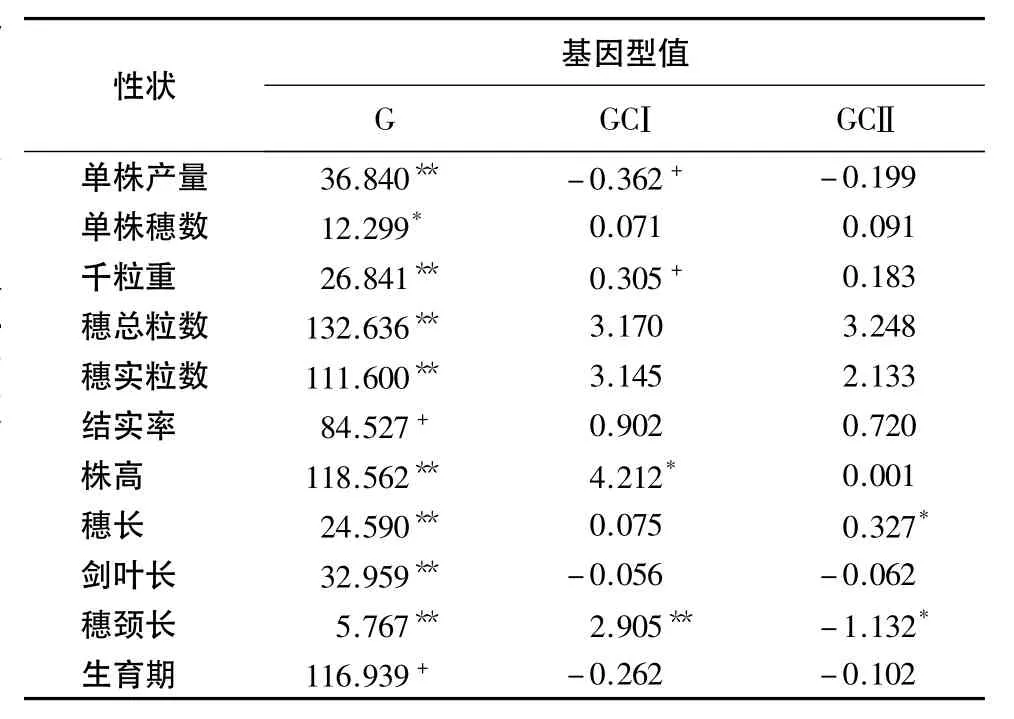
表3 不育胞质杂种(AR)和可育胞质杂种(BR)的F1平均基因型值的预测1)Table 3 Mean prediction of F1genotype value of traits in two cross types
2.5 不育细胞质的核质互作效应对CMS-FA胞质杂交稻F1产量相关性状的杂种优势的影响
由表4可知,CMS-FA胞质杂种除单株穗数和生育期外的其他所有性状,核基因效应(G)所产生的正向群体平均优势均达显著或极显著的水平,表明水稻新质源(CMS-FA)遗传群体F1在这些性状上具有明显的群体平均优势,且以穗颈长最高(1.062),其次为单株产量(0.289)、每穗实粒数(0.149)、株高(0.133)、千粒重(0.088)、每穗总粒数(0.077)、结实率(0.056)和穗长(0.044),剑叶长最小(0.038).核质互作基因效应(GC)所产生的群体平均优势,除穗颈长外其他性状总的程度均明显小于核基因(G)的作用,但在单株产量、千粒重、每穗实粒数、株高、穗长和穗颈长等方面仍显著存在.不育胞质和可育胞质与核基因的互作,均使千粒重表现为显著的正向优势,而单株产量表现为极显著的负向优势;不育胞质×核基因互作使每穗实粒数、株高和穗颈长产生显著的正向优势,而可育胞质×核基因互作使穗长显著增长以及穗颈长显著缩短.
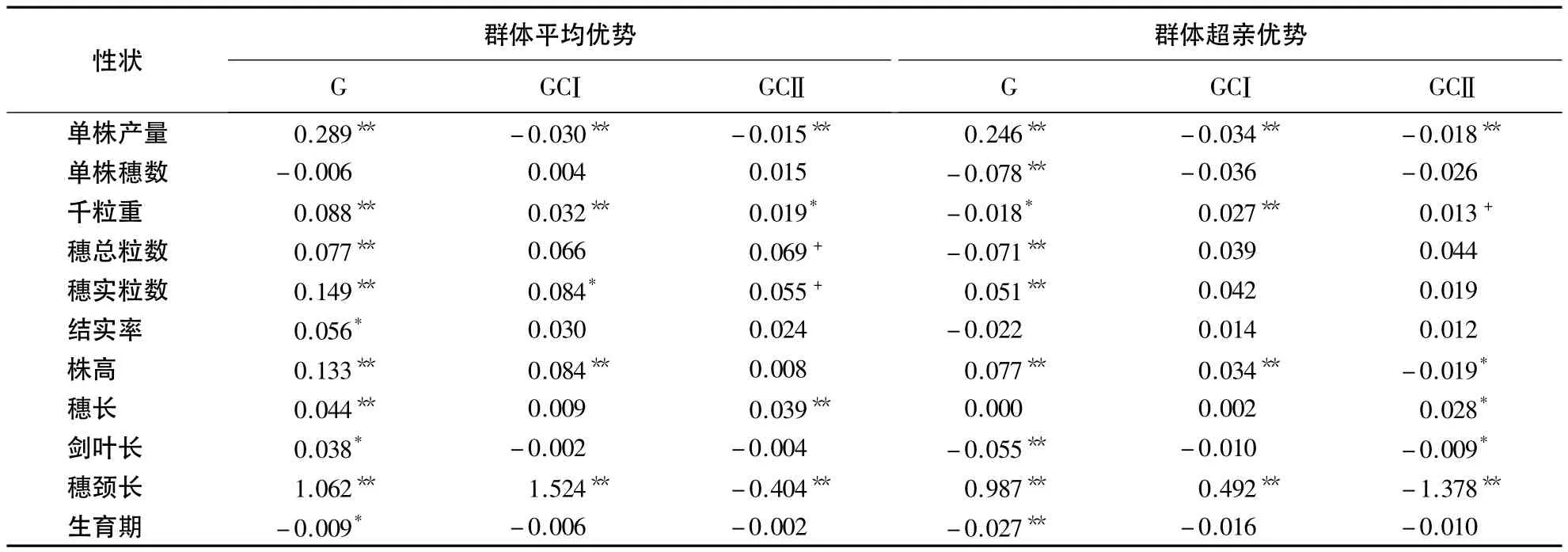
表4 不育胞质杂种(AR)和可育胞质杂种(BR)的群体杂种优势表现1)Table 4 Heterosis based on population of traits in two cross types
核基因效应(G)在单株产量、每穗实粒数、株高和穗颈长等4个性状上所产生的群体超亲优势(HBP)表现为极显著的正向优势,且以穗颈长最高(0.987),其次为单株产量(0.246)、株高(0.077)和每穗实粒数(0.051).在单株穗数、千粒重、每穗总粒数、剑叶长和生育期等5个性状上表现为显著或极显著负向优势,以单株穗数最大(-0.078),其次为每穗总粒数(-0.071),千粒重最小(-0.018).不育胞质×核基因互作所产生的群体超亲优势(HBP)使千粒重、株高和穗颈长表现为显著的正向优势,但却使单株产量显著降低;可育胞质×核基因互作使单株产量显著降低、株高、剑叶长和穗颈长缩短.
3 讨论
福建农林大学新质源杂交稻育种技术研究课题组研发出的新型CMS-FA胞质杂交稻[4-9],对增加我国水稻籼型三系不育系胞质类型和丰富杂交稻细胞质的遗传多样性有重要意义.水稻不育细胞质对杂种F1产量相关性状有一定影响[17-22],通过对不育胞质遗传效应的研究,可以评价现有和新发现的不育系细胞质效应的优劣,选择可恢复性好的细胞质不育系,或者选育恢复力强的恢复系与之配组,均可达到基本克服或最大限度减轻不育细胞质对杂交稻产量等相关性状的负效应[1,23].因此,评价CMS-FA胞质杂交稻不育胞质的遗传效应及其对杂种F1产量相关性状的影响,对CMS-FA胞质杂交稻应用有重要作用.
国内外已有较多关于水稻雄性不育细胞质对籼型杂交稻或粳型杂交稻遗传效应的研究报道[1,16-30],但结果不完全相同.有较多研究结果表明,不育细胞质在杂交稻F1的株高、每穗实粒数、结实率和单株产量等性状上表现出明显的负效应[1,17-20,23-26],但不同研究在抽穗期、单株穗数、每穗总粒数及千粒重等性状上的结果存在较大差异[1,18-21,23,26].本研究结果表明,CMS-FA 胞质杂交稻不育细胞质效应主要表现在穗颈长、千粒重和生育期等性状上,对其它8个性状的影响不明显,并且多数性状对提高产量有增强效应的趋势.这与前人的研究结果不完全相同,可能是因为CMS-FA胞质杂交稻不育系可恢复性好,选育的恢复系恢复力强,在一定程度上消除了不育胞质中不育性的影响.
在本研究中,虽然细胞质对穗颈长的负效应达到极显著水平,但对CMS-FA胞质杂交稻产量没有明显影响,不会成为CMS-FA胞质杂交稻杂种优势利用的主要障碍.因此,CMS-FA胞质杂交稻不育细胞质没有不良的细胞质效应.2010年通过福建省审定的CMS-FA胞质杂交稻组合金农2优3号在区试中的表现好、产量高,两年区试平均产量达到488.74 kg,比对照汕优63平均增产9.025%,均达极显著水平,米质达部颁三等优质食用稻品种标准,这一结果也证实了CMS-FA胞质杂交稻不育细胞质可作为杂交稻育种的优良不育胞质利用.
本研究根据盛孝邦介绍的方法[10]分析CMS-FA胞质杂交稻不育细胞质的遗传效应,但此方法无法估算核质互作效应[28,30].因此采用数量性状的加性、显性(核基因)与核质互作模型及其统计分析方法[11-16],检测和分析CMS-FA胞质杂交稻核质互作效应及其对杂种产量相关性状的影响.结果表明,核质互作效应对CMS-FA胞质杂交稻F1的单株穗数、千粒重、每穗总粒数、结实率、株高、穗长、穗颈长和生育期等性状有明显的作用,但与核基因效应相比,其影响程度较小.不育胞质与核基因互作的结果会导致一定程度的株高增加和穗颈长增长,但对CMS-FA胞质杂交稻杂种优势利用的影响较小.
[1]孙叶,顾燕娟,张宏根,等.水稻3种不育细胞质遗传效应的比较研究[J].扬州大学学报:农业与生命科学版,2006,27(2):1-4.
[2]曾千春,周开达,朱祯,等.中国杂种优势利用现状[J].中国水稻科学,2000,14(2):243-246.
[3]程式华,李建.现代中国水稻[M].北京:金盾出版社,2007:208-237.
[4]王乃元.野生稻(O.rufipogon)新胞质改良不育系稻米品质的研究[J].作物学报,2006,32(2):253-259.
[5]王乃元.野生稻(O.rufipogon)雄性不育恢复系的研究[J].作物学报,2006,32(12):1884 -1891.
[6]王乃元,梁康迳,李毓,等.新质源(CMS-FA)杂交稻系统的亲本资源筛选[J].作物学报,2008,34(9):1549-1556.
[7]王乃元,梁康迳,李毓,等.水稻新质源(CMS-FA)雄性不育恢复基因的遗传[J].作物学报,2008,34(11):1929-1937.
[8]王乃元.新质源(CMS-FA)杂交稻育种技术探索与研究[J].中国农业科技导报,2009,11(S2):7-12.
[9]王洪飞,王乃元,李毓,等.新质源(CMS-FA)杂交稻产量相关性状的遗传效应与杂种优势分析[J].中国农业科学,2010,43(2):230-239.
[10]盛孝邦.杂交水稻细胞质对农艺性状遗传效应的研究[J].中国水稻科学,1987,1(3):155-170.
[11]朱军.遗传模型分析方法[M].北京:中国农业出版社,1997:98-111,240-255.
[12]朱军.作物杂种后代基因型值和杂种优势的预测方法[J].生物数学学报,1993,8(1):32-44.
[13]朱军,季道藩,许馥华.作物品种间杂种优势遗传分析的新方法[J].遗传学报,1993,20(3):262-271.
[14]ZHU J,WEIR B S.Diallel analysis for sex-linked and maternal effects[J].Theoretical and Applied Genetics,1996,92(1):1-9.
[15]朱军.广义遗传模型与数量遗传分析新方法[J].浙江农业大学学报,1994,20(6):551-559.
[16]沈圣泉,薛庆中.不同细胞质的核质互作对籼粳杂种F1代主要农艺性状的影响[J].中国水稻科学,1997,11(1):6-10.
[17]朱英国.水稻不同细胞质类型雄性不育系的研究[J].作物学报,1979,5(4):29-38.
[18]杨仁崔,刘抗美,卢浩然.水稻“野败”不育胞质对杂种一代的影响[J].福建农学院学报,1980,1(2):14-21.
[19]杨仁崔,刘抗美,卢浩然.水稻冈型不育细胞质对杂种一代的影响[J].中国农业科学,1984,17(3):1-5.
[20]盛孝邦.水稻不育系胞质遗传效应的初步研究Ⅰ.不育系胞质对杂交水稻农艺性状的影响[J].华中农学院学报,1982,1(3):1 -12.
[21]广西农科院水稻杂优组.水稻雄性不育细胞质对子一代主要性状的影响[J].中国农业科学,1982,15(4):7-12.
[22]邢少辰.籼型杂交水稻不育细胞质对杂种一代主要农艺性状的影响[J].广西农学院学报,1990,9(3):15-22
[23]汤述翥,张亚东,孙红芹,等.水稻同核异质广亲和不育系细胞质效应的研究[J].作物学报,2003,29(2):202-207.
[24]陈银辉,蔡俊迈,卢浩然.水稻雄性不育细胞质和核质互作的遗传表现[J].福建农学院学报,1987,16(3):190-197.
[25]王才林,汤玉庚.杂交粳稻不育细胞质遗传效应的研究[J].作物学报,1990,16(4):335-341.
[26]王才林,汤述翥,汤玉庚.杂交粳稻同核异质不育系细胞质效应的研究[J].中国水稻科学,1998,12(2):65-71.
[27]王文明,周开达,文宏灿,等.胞质效应在杂交水稻主要数量性状上的多样性[J].中国水稻科学,1997,11(2):65-69.
[28]王彦荣,王妍,代贵金,等.水稻雄性不育细胞质效应研究综述[J].辽宁农业科学,2000(3):33-36.
[29]牟凤娟,张树华.水稻CMS不育系细胞质遗传效应的研究进展[J].植物遗传资源科学,2002,3(3):47-52.
[30]仇秀丽,王洪飞,李毓,等.杂交水稻细胞质遗传效应的研究进展[J].云南农业大学学报,2009,24(3):455-458.
