味觉厌恶学习对大鼠舌乳头上皮leptin表达的影响
2013-09-21李海艳程振宇
李海艳,程振宇
(承德医学院,河北承德 067000)
动物摄取某种新异的味觉刺激后,若出现恶心、呕吐、腹泻等内脏不适,动物便可“记住”这种食物的味觉特征,减少或拒绝摄取具有相同味觉特征的食物,这种行为的变化过程称为条件性味觉厌恶学习(CTA),是动物避免重复摄取有毒物质的一种重要学习机制。味觉对于调节摄食,控制体重起着重要作用。脂肪细胞产生的瘦素(leptin)能够调整食物的吸收、能量的消耗和体重。目前认为,味觉器官是leptin作用于外周的靶器官之一,leptin通过与味觉细胞的ob-Rb受体结合抑制机体对甜味物质的反应,降低味觉感受器的敏感性。因此,由leptin引起的味觉改变可能参与了对食物吸收的调整[1]。本研究通过建立大鼠CTA模型,采用免疫组化染色法观察了CTA模型大鼠舌乳头上皮leptin表达的变化。
1 材料与方法
1.1 实验动物分组 清洁级成年雄性SD大鼠20只,体重250-300g,购自河北医科大学实验动物中心(许可证号:SCXK(冀)2008-1-003)。大鼠随机分为CTA组和对照组,每组10只大鼠,两组均进行口腔插管手术,建立模型时CTA组腹腔注射LiCl(20ml/kg)、对照组腹腔注射生理盐水(20ml/kg)。
1.2 CTA模型的建立
1.2.1 口腔插管手术:大鼠腹腔注射0.5%戊巴比妥钠(45mg/kg)麻醉,固定于脑立体定位仪上,暴露顶骨。将医用单孔聚乙烯管的一端,由皮肤切口处经耳眼间皮下组织导入大鼠颊部并穿入口腔,在口腔内加一垫圈固定;在顶骨表面固定两个螺丝钉,将导管上端用牙托粉固定于螺丝钉上,伤口处洒上灭菌结晶磺胺消炎。
1.2.2 建立模型:口腔插管手术后3-5d,经口腔插管向口腔内注入0.2mol/L蔗糖溶液5ml,5min内注完;20min后,腹腔注射0.15mol/L的LiCl(20ml/kg)。禁食禁水12h后,用两瓶法测定水和蔗糖的摄取量。如果对蔗糖的摄取量小于0.5ml,说明建立了一次CTA,重复建立3次予以巩固。
1.3 免疫组织化学染色法检测leptin的表达 造模成功后,大鼠腹腔注射25%氨基甲酸乙酯(2g/kg)麻醉,打开胸腔暴露心脏,经升主动脉插管,150ml生理盐水快速灌流冲洗,然后用4%多聚甲醛灌流固定1.5h,取舌,入4%多聚甲醛后固定4-6h,常规石蜡包埋,连续切片,片厚5μm。采用SP免疫组织化学染色法观察大鼠舌leptin的表达,兔抗leptin多克隆抗体(北京中杉金桥生物技术有限公司)1:300稀释,DAB(美国Sigma公司)显色,苏木素复染。阴性对照以PBS代替一抗。
1.4 切片观察和细胞计数 每只大鼠观察5张切片,每张切片在200倍显微镜下取6个视野,Image-Pro Plus(IPP)分析软件系统自动计数两种舌乳头上皮限定面积(丝状乳头和菌状乳头,270μm×250μm)内阳性细胞的数目,换算为单位面积内leptin阳性细胞数(/mm2),取平均值作为该舌乳头上皮leptin蛋白表达的相对水平。
1.5 统计学处理 采用SPSS 14.0统计软件处理数据,两样本均数的比较采用t检验,P<0.05为差异有统计学意义。
2 结果
丝状乳头、菌状乳头和轮廓乳头上皮均可见leptin免疫反应阳性细胞,leptin阳性产物位于细胞质,呈棕黄色。与对照组比较,CTA组大鼠舌乳头上皮的染色明显加深,且CTA组大鼠舌丝状乳头、菌状乳头上皮leptin的表达水平明显高于对照组,差异具有统计学意义(P<0.01)。见附表。
附表 各组大鼠舌乳头上皮leptin的表达(±s,单位:细胞数/mm2)
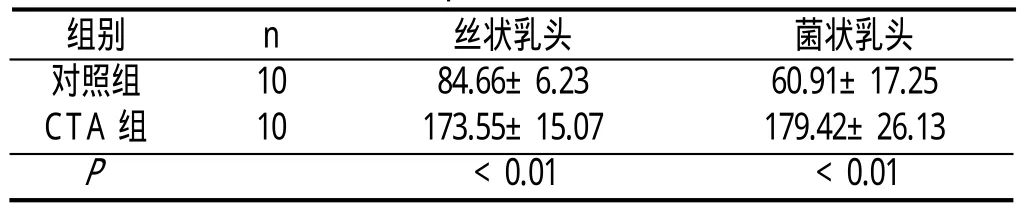
附表 各组大鼠舌乳头上皮leptin的表达(±s,单位:细胞数/mm2)
组别 n 丝状乳头 菌状乳头对照组 10 84.66±6.23 60.91±17.25 CTA组 10 173.55±15.07 179.42±26.13 P <0.01 <0.01
3 讨论
leptin是肥胖基因(ob基因)的蛋白产物,主要在脂肪组织中表达,被认为是一种饱食信号,主要调节能量平衡和控制体重。向遗传性肥胖小鼠(ob小鼠)的脑脊液注入leptin后,小鼠的进食量明显减少,体质量也逐渐下降[2]。Monteleone等[3]和Waelput等[4]发现,leptin具有广泛的生物学效应:一是通过作用于下丘脑的摄食中枢产生饱胀感,从而抑制摄食行为;二是作用于其它组织的leptin受体,包括肝、脾、肺、肾、心、胰腺等,直接或间接发挥生物学效应[5]。目前,有研究认为leptin也可作用于味觉器官。
动物建立味觉厌恶学习后,有恶心、呕吐、腹泻等内脏不适的表现,对摄食的适应性调节能力降低,进而导致胃排空的延迟。leptin水平及其受体表达的多少和性状的改变,可以改变胃的顺应性,抑制摄食,延迟胃排空。原因是leptin可直接激活迷走神经传入神经元,刺激信号通过迷走神经到达孤束核(NTS)刺激leptin敏感神经元,再作用于下丘脑外侧区(LH)、下丘脑腹内侧区(VMH)及弓状核(ARC),抑制胃的运动和摄食,因此提示leptin能够调节摄食、调节胃排空[6]。近年有研究报道,leptin还能影响小鼠的记忆和学习活动。敲除leptin受体基因的db/db小鼠,依然能建立条件性味觉厌恶学习,但却加快了CTA的消退过程,原因可能是leptin受体差异性地分布于CTA相关脑区[7]。另外,leptin可能通过杏仁核、海马、皮质、臂旁核的leptin受体发挥作用,而且leptin受体在CTA大鼠脑内神经元中表现出内移现象,表明在CTA的形成和维持中,leptin受体的活性或功能发生了变化,提示leptin在味觉和内脏信息的相互作用中具有重要作用[8]。而CTA也是味觉信息和内脏信息发生整合作用的结果,动物建立CTA后是否会影响外周器官—舌leptin的表达,目前尚未见相关报道。
本研究发现,大鼠丝状乳头、菌状乳头及轮廓乳头上皮均可见leptin表达,并且CTA大鼠舌丝状乳头、菌状乳头上皮leptin的表达水平明显高于对照组大鼠;因此提示CTA影响了leptin蛋白在舌乳头上皮的表达,进一步阐明了CTA和leptin蛋白的关系,充实了CTA理论,但相关机制有待于深入研究。
[1]Shigemura N,Ohta R,Kusakabe Y, et al. Leptin modulate behavioral responses to sweet substances by influencing periphera taste structures[J].Endocrinology,2004,145(2):839-847.
[2]王田.瘦素的研究进展[J].现代中西医结合杂志,2009 18(28):3525-2526.
[3]Monteleone P, DiLieot A,Castaldo E, et al. Leptin functioning i eating disorders[J]. CNS spectr, 2004, 9(7): 523-529.
[4]Waelput W, Brouckaert P, Broekaert D, et al. A role for lepti in the systemic inflammatory response syndrome(SIRS) an in immune response [J].Curr Drug Targets Inflamm Allergy 2002,1(3):277-289.
[5]李久红.对瘦素的进一步认识[J].武警医学院学报,2008 17(1):76-78,82.
[6]于卫星,魏良洲.瘦素与胃排空[J].国际消化病杂志,2007,27(6):428-430.
[7] Ohta R, Shigemura N,Sasamoto K, et al. Conditioned taste aversion learning in leptin-receptor-deficient db/db mice[J]. Neurobiol Learn Mem,2003,80(2):105-112.
[8]韩蓁,闫剑群,宋新爱,等.瘦素受体在条件性厌味大鼠脑的表达[J].第四军医大学学报,2003,24(1):11-13.
