长白山杨桦次生林主要树种的空间分布格局及其关联性1)
2013-09-18亢新刚孟京辉郭韦韦
杜 志 亢新刚 孟京辉 孔 雷 郭韦韦 岳 刚
(省部共建森林培育与保护教育部重点实验室(北京林业大学),北京,100083)
种群空间结构与不同的进程和外力密切相关,包括种间和种内竞争、种子散布、干扰、环境异质性等[1-3],植物空间结构的分析是研究种群特征、种群和环境相关性的重要方法[4],有助于揭露群落演替的潜在进程和机制。近年来,点格局统计在生态学中得到广泛运用[2,5-8],作为研究不同空间尺度上空间模式的有效工具,它克服了传统方法只能在单一尺度上研究物种空间分布和联系的不足。
中国次生林的面积已经占了森林面积的一半以上[9],东北是中国木材生产的主产区,由于长期的过伐和缺乏保护,大量原始林遭受到了严重破坏而转换为次生林,杨桦林在东北次生林占相当大的比重,成为重要的后备森林资源[10]。为了可持续经营此类型森林,了解其树种的生物特征和潜在生态进程十分关键。国内学者对于东北杨桦次生林进行了大量研究,主要集中在物种组成和直径结构分布[11-12]、种群性比和分布情况[13-14]、生长过程和演替动向[15-16]、林隙更新[17-18]等方面。但是,关于杨桦次生林主要树种的空间分布模式和空间关联性的研究很少,尤其是不同林层的空间结构的分析。笔者运用O-ring统计分析1 hm2杨桦次生林调查数据,旨在认识其空间格局和关联性,为次生林的正向演替提供科学依据,为将来的森林群落恢复和重建工作打下基础。
1 研究地概况
研究地位于中国东北汪清金钩岭林场长白山的杨桦次生林(43°22'N,130°15'E)。次生林位于林场海拔300~1200 m处,受海洋湿气团的影响,本区具有温带海洋季风气候的特点。月均温3.9℃,积温10℃以上的将近2473℃。年降水量为500~600 mm,无霜期为110~130 d,土壤为棕灰化土。本研究区的乔木树种有:白桦(Betula platyphylla)、椴树(Tilia amurensis)、枫桦(Betula costata)、红松(Pinus koraiensis)、云杉(Picea jezoensis)、冷杉(Abies nephrolepis)、山桃(Amygdalus daridiana)、山杨(Populus davidiana)、蒙古栎(Quercus mongolica)、色木(Acer mono)、水曲柳 (Fraxinus mandcshurica)、落叶松(Larix olgensis)、大青杨(Populus ussuriensis)、暴马丁香(Syringa amurensis)等。
2 研究方法
2.1 样地布设与调查
于2011年夏在金钩岭林场建立1 hm2的杨桦次生林样地。该样地远离公路,近期人为干扰较少。研究区域依次划分为25个20 m×20 m次级样地,在每个次级样地中,对每个胸径大于5 cm的林木进行定位,记录树名,量测胸径、树高、冠幅、第一枝下高。
根据样地的树种重要值,以及树种在演替过程中的重要性,并结合前人在长白山杨桦次生林中的研究[11,19-20],选取白桦、枫桦、红松和冷杉 4 个主要树种作为研究对象。为分析森林的空间结构,调查者常将林分划分为3或4个高度层[5,21],树木的高度数据通过运用理查德生长方程[22]来推导。先将4个树种划分为3个高度级:h<10 m、10 m<h≤20 m、h≥20 m,但发现高于20 m的林木在次生林中相当少。所以,本研究将所有的树种划分为2个高度级:h<10 m和h≥10 m,并用L和U分别表示;分析时也将同一树种视为一个整体,不进行分级,用A表示。
2.2 空间结构分析方法
点格局统计中的二阶函数被视为描述树种密度变化的最有效的函数[2,23]。其中,二阶点格局分析Ripley’s K 得到了广泛应用[6-7,24-26],它能描述绘图区域内点对点的距离,为监测不同种群和模式尺度提供了可能性[2,25]。但是,Ripley’s K 函数在分析树种空间格局时存在一定的缺陷[23,25],其分析是基于半径为r,圆心为指定点的圆。在大尺度上进行分析时,会统计小尺度部分引起积累效应,而影响最终结果。
O-ring统计从Ripley’s K函数发展而来,用圆环取代Ripley’s K函数的圆,分离了特定的距离等级,有效地消除积累效应,能分析任意给定距离的空间格局[25]。单变量O-ring统计[O(r)]可分析单一目标(如一个树种)的空间分布格局,双变量O-ring统计[O12(r)]可分析两个对象(如两个树种)的空间关联性。笔者运用前者来分析主要树种在不同林层的空间分布格局,运用后者来研究种内种间关联性。
单变量统计对空间分布格局分析中,空间异质性的存在将导致分布格局并非由树种相互作用形成,从而易导致格局误判,因此对于零模型(null model)的准确选择极为重要[2]。结合样地树种的空间分布图,若树种表现出明显空间异质性,则使用异质性Possion过程模型;反之,则采用完全空间随机过程模型。对于双变量统计,假定高林层的林木对低林层的林木有压制作用,而低林层的林木对高林层的林木影响不大[21,27],然后根据前提条件零假设模型分析树种相关性。用Monte Carlo模拟结果,若单变量O(r)值(双变量O12(r)值)在上包迹线以上,则为聚集分布(空间上正关联);若在下包迹线以下,则为均匀分布(空间上负关联);若在上下包迹线之间,表示随机分布(空间上无相关,相互独立)。
运用 R Spatstat软件包(2.14.0版)绘制树种分布图,生态学软件Programita(2008版)完成O-ring统计分析。采用空间尺度0~50 m,根据相应零假设模型,进行19次Monte Carlo模拟得到95%的置信区间[28]。
3 结果与分析
3.1 林分结构
如表1所示,植株数量最多为白桦,670株,胸高断面积为11.41 m2·hm-1,显然白桦是本样地最主要的树种。色木、落叶松、云杉和大青杨的基本结构特征统计值(如植株数量、胸高断面积和重要值)都低于其它树种。白桦的重要值为42.12,明显高于其它树种;红松的重要值排第二,为20.64,枫桦为6.64。冷杉的重要值高于山桃,分别为6.13和5.99。山杨的断面积较大,植株数量很少,而表现出较小的重要值。树种结构特征数据说明此样地为次生林,白桦、红松、枫桦和冷杉为主要树种。

表1 长白山杨桦次生林1 hm2样地的树种基本结构特征
表2显示了整个样地以2 cm为径阶划分的直径结构。白桦的直径分布出现了两个高峰值,即8、16 cm处,植株数量明显多于其它树种,中等径阶的植株分布较多,说明样地前期为阳性树种提供了充足的光照条件。红松和枫桦的直径分布在20 cm以内,显示出明显的递减分布现象,并且直径大于20 cm的植株数量相当少。冷杉为的株数在8 cm处最多。表明先锋树种白桦侵入后,林分开始郁闭,为顶级耐阴树种提供了生长条件,针叶树种开始生长,起测径阶附近植株数量最多,且随径阶增大而减少。
3.2 不同林层主要树种的空间分布
不同林层中4个树种的分布模式如图1所示。3个林层中的白桦均显示均匀或随机分布,低林层中的红松同样如此(图1b)。样地中央区域枫桦分布少(图1c),冷杉主要分布在样地南部(图1d),表明其聚集分布格局。各林层树种密度差异较大,高度小于20 m的林木株数要远远多样高于20 m的。
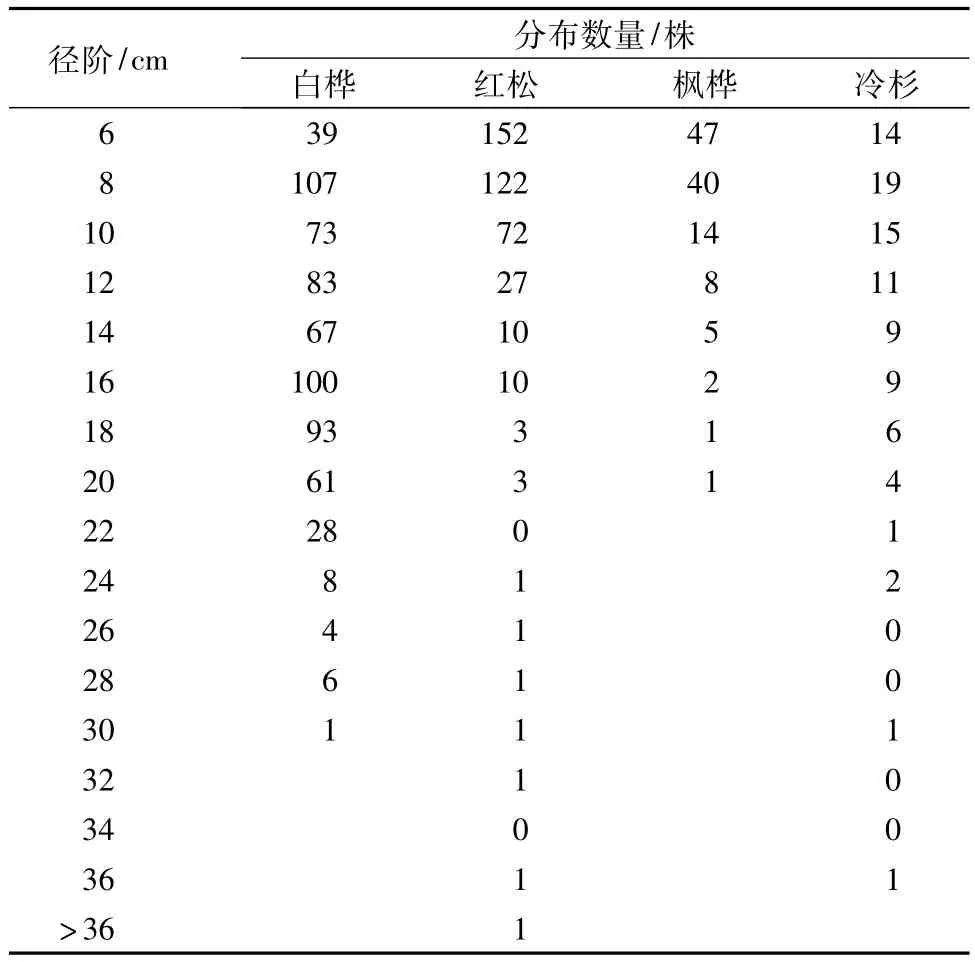
表2 杨桦次生林1 hm2样地的主要树种径阶分布数量
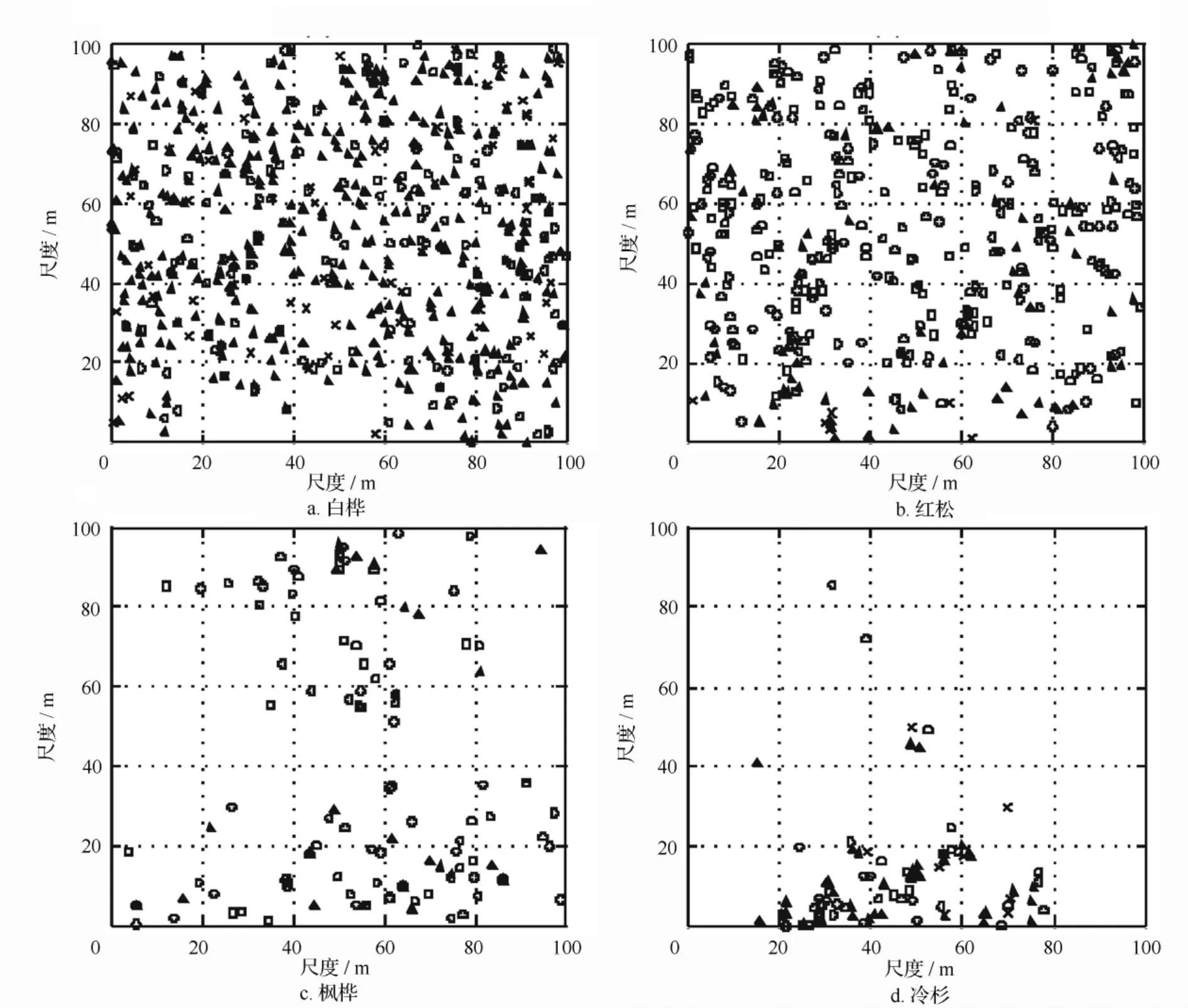
图1 主要树种不同高度级的空间分布
3.3 主要树种在不同林层的空间分布格局
各个树种在不同林层和尺度上的空间分布格局多样(图2)。整个白桦种群在18~21 m和18~32 m范围内显示聚集分布(图2a)以外,在大多范围内都表现为明显的随机分布。高林层白桦与低林层白桦的分布格局比较,高林层中部分尺度为聚集分布(图2b),低林层中白桦随机分布占主导(图2c)。整个红松种群和高林层红松随机分布普遍,在较大尺度上,高林层红松出现均匀分布,低林层的红松在空间尺度≤6 m范围内聚集分布显著,在12~44 m范围内空间分布趋向于聚集。整个枫桦种群和低林层枫桦≤24 m范围上呈现聚集分布,继而随尺度增大向随机分布、均匀分布变化(图2g和2i),而高林层枫桦则在整个尺度上呈现随机分布。整个冷杉种群以及高低林层的冷杉都在≤30 m尺度上表现聚集分布,并随尺度增加聚集度降低而转变为随机分布(图2j和 2k、2l)。
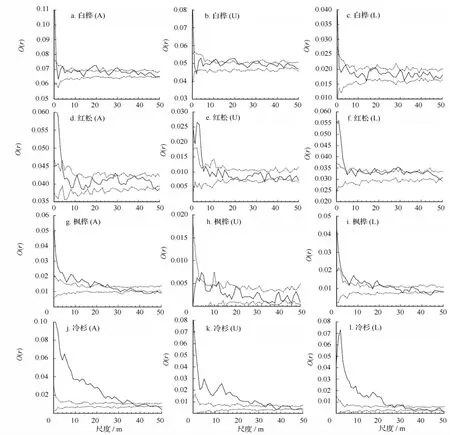
图2 主要树种在不同林层中的空间分布格局
不同树种在高低林层中的分布格局存在差异,低林层的红松和枫桦在部分尺度上表现为较明显的聚集分布,处于高林层的树随机分布普遍,而高林层的白桦则出现了聚集分布,而低林层白桦完全表现为随机分布。
3.4 不同林层中的种内和种间空间关联性
表3说明低林层白桦与高林层的林木在大多尺度上表现为相互独立,与高林层红松、冷杉在大尺度上表现为负相关。低林层的红松与高林层的4个树种表现的关联性各异:①与白桦在小尺度和大尺度上基本为正相关;②与红松<10 m尺度上基本表现正相关,其它尺度上以无相关为主;③与枫桦在大多尺度上表现为相互独立;④在<14 m范围内与冷杉成负相关,随尺度增加逐渐表现为无相关。低林层枫桦与高林层白桦、红松主要表现为无相关性,而与高林层枫桦、冷杉在<25 m尺度上大多呈现正相关,并随尺度增加转变为无相关。低林层的冷杉与高林层白桦在2~25 m范围内呈相互排斥的关联性,与高林层的红松1~10 m范围内,以及与高林层冷杉在1~25 m范围内表现为明显正相关,与高林层枫桦在所有尺度上表现相互独立。

表3 低林层和高林层的树种种内种间关联性

续(表3)
低林层中分析树种间的空间关联性。白桦与红松、枫桦基本表现为相互独立,而白桦与冷杉、红松与冷杉在所有尺度上相互排斥的现象普遍;红松与枫桦则在尺度范围15 m内呈现负相关,>15 m后关联性向无相关变化。枫桦和冷杉在<15 m范围内表现正相关。高林层两树种关联性中,15 m以内的小尺度上,白桦和红松、白桦和冷杉、红松和冷杉都呈现负相关,白桦和枫桦、红松和枫桦之间无相关占主导,枫桦和冷杉之间以正相关为主;尺度>15 m后,随尺度增加高林层树种两两间的关联性有向相互独立变化的趋势。
4 结论与讨论
研究地为异龄次生林,具有丰富的树种和复杂的林分结构(表1)。先锋强阳性树种如白桦,占样地植株数量的大部分,同时表1显示样地各树种的平均胸径较小,这都明显说明该样地处于演替的初级阶段。然而,该样地有不少红松,且红松最大胸径超过50 cm,平均胸径将近10 cm,这暗示着该样地所在林分由阔叶红松林遭受采伐和严重破坏后转换而来,此推断与先前森林经营记录情况相吻合[29]。
经营措施造成的林分环境差异,种群生物学特性的不同,以及林分范围上的资源分配不均都是导致空间分布格局多样的原因[1-2]。受到严重破坏的森林,林中往往形成若干大小不一的林窗,林窗内光照充足,有利于强阳性的先锋种快速生长,从而聚集分布在林分中,论证了图2b中高林层白桦一定尺度上聚集分布的表现。当先锋种白桦生长起来,以及未采伐的红松等树种,占据高林层而使林地郁闭,从而抑制下层强阳性树种的生长更新,造成了低林层的白桦基本表现为随机分布(图2c)。
通常,随着林木的生长,对光照、养分、水分、生长空间的需求增大,而有限的资源无法满足所有林木的生长,种内与种间竞争加剧,个体死亡率上升,自然稀疏效应产生,从而导致种群分布格局由聚集趋向于随机[5,30-31]。本研究得到了一致的结果,高林层的红松(图2e)和枫桦(图2h)在大多尺度上都呈现随机分布。红松球果大且重,无法借助风力或重力传播[32],其散布大多依赖动物的活动[33],种子有限的散布特性是低林层红松(图2l)在一定尺度上呈现聚集分布的原因。在演替阶段中,枫桦、冷杉在白桦侵入后生长出来,表1中枫桦、冷杉植株数量也较少,说明此两树种在林分里开始繁殖的时间不长,远未达到充分利用空间资源的程度,且属于耐阴树种,所以图2中的枫桦、冷杉在大部分尺度上表现为聚集分布。利用同一树种在不同生长阶段下表现出的空间分布规律,即可采取相应的经营措施,若种植红松幼苗可考虑在阴处密植,而对红松大树的栽植则应保证具有充足光照,足够大的生长空间。
树种间空间关联性中(表3),低林层的红松与高林层白桦、红松在小尺度上都呈正相关,反应了红松幼年较耐阴和种子散布能力有限的特点;而与高林层冷杉主要表现为负相关,主要是因为冷杉凋落的针叶对红松幼苗的生长有不利影响[34],所以对于红松的补植应有选择地在红松、白桦大树下进行,避免在冷杉树下栽植。随着红松年龄增加,耐阴性逐渐减弱,作为林分的顶级树种,逐渐在环境资源争夺中占据主动,与其它树种相互依赖性减弱而表现为负相关,文中高林层的红松和白桦、红松与冷杉反映了这种情况。顶级树种红松和冷杉与先锋树种白桦在各林层上都表现出负相关,尤其高林层中相互排斥现象明显,表现了不同演替阶段的树种间的激烈竞争,暗示了样地暗针叶林正向演替的趋势。若经营的目的树种为经济价值高的红松,可考虑人为采伐一些红松大树旁边的白桦,为红松提供更多的阳光、养分等生长空间,促进其生长。
随着空间尺度的增加,各树种的空间分布在较大尺度上都表现为随机分布。另外,种间关联性也随着空间尺度变化而变化,分别分析高低林层树种关联性时,不同树种间的正相关或负相关主要发生在小尺度范围内,关联性随尺度增大而减弱,趋于不相关,这反应了空间格局和关联性对尺度的依赖性,此结果与前人研究结论[35]一致,不同尺度上表现的不同空间分布和关联性涉及到许多潜在的、复杂的生物学特征(如繁殖特征、树冠生长、耐阴程度等)和生态学过程(如光照状况、土壤理性、湿度情况等),这些仍需深入研究。
杨桦次生林处于森林演替的初级阶段,具有巨大潜在的经济价值,了解其林分结构,空间分布格局和关联性,能为森林经营提供一定的指导,在发挥效益的同时也有助于引导森林的正向演替。将来应再设立代表不同演替阶段的大样地,进行对比分析,并结合不同空间结构下林木的生长状况,根据规律探讨不同树种的抚育采伐方案,确定具体的森林经营工作方案。
[1]Condit R,Ashton P S,Baker P,et al.Spatial patterns in the distribution of tropical tree species[J].Science,2000,288:1414 -1418.
[2]Stoyan D,Penttinen A.Recent applications of point process methods in forestry statistics[J].Statistical Science,2000,15(1):61-78.
[3]张金屯,孟东平.芦芽山华北落叶松林不同龄级立木的点格局分析[J].生态学报,2004,24(1):35-40.
[4]Watt A S.Pattern and process in the plant community[J].British Ecological Society,1947,35(1/2):1 -22.
[5]张健,郝占庆,宋波,等.长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性[J].应用生态学报,2007,18(8):1681-1687.
[6]张赟,张春雨,赵秀海,等.长白山次生林乔木树种空间分布格局[J].生态学杂志,2008,27(10):1639-1646.
[7]陈列,赵秀海,张赟.长白山北坡椴树红松林空间分布及其空间关联[J].北京林业大学学报.2009,31(3):6-10.
[8]杜志,亢新刚,包昱君,等.长白山云冷杉林不同演替阶段的树种空间分布格局及其关联性[J].北京林业大学学报,2012(2):14-19.
[9]李国猷.北方次生林经营[M].北京:中国林业出版社,1992:1-5.
[10]吉林森林编辑委员会.吉林森林[M].吉林:吉林省科学技术出版社;北京:中国林业出版社,1988:231-245.
[11]郝占庆,张健,李步杭,等.长白山次生杨桦林样地:物种组成与群落结构[J].植物生态学报,2008,32(2):251-261.
[12]龚直文,亢新刚,杨华,等.长白山杨桦次生林直径结构研究[J].西北林学院学报,2009,24(3):1-6.
[13]张春雨,王德胜,魏彦波,等.山杨、水曲柳种群密度与种群性比的关系[J].林业科学,2010,46(7):16-21.
[14]张春雨,魏彦波,王德胜,等.水曲柳种群性比及空间分布[J].林业科学,2010,46(10):167-172.
[15]龚直文,亢新刚,顾丽,等.长白山杨桦次生林生长过程与演替动向分析[J].林业科学研究,2009,22(3):379-384.
[16]赵伟,金慧,李江楠,等.长白山北坡天然次生杨桦林群落演替状态[J].东北林业大学学报,2010,38(12):1-3.
[17]宋新章,李冬生,肖文发,等.长白山区次生阔叶林采伐林隙更新研究[J].林业科学研究,2007,20(3):302-306.
[18]宋新章,张智婷,肖文发,等.长白山次生杨桦林采伐林隙乔灌木幼苗更新比较研究[J].林业科学研究,2008,21(3):289-294.
[19]赵淑清,方精云,宗占江,等.长白山北坡植物群落组成、结构及物种多样性的垂直分布[J].生物多样性,2004,12(1):164-173.
[20]龚直文,顾丽,亢新刚,等.长白山森林次生演替过程中林木空间格局研究[J].北京林业大学学报,2010,32(2):92-99.
[21]Hao Z Q,Zhang J,Song B,et al.Vertical structure and spatial associations of dominant tree species in an old-growth temperate forest[J].Forest Ecology And Management,2007,252:1 -11.
[22]赵俊卉,亢新刚,张慧东,等.长白山3个主要针叶树种的标准树高曲线[J].林业科学,2010,46(10):191-194.
[23]George L,Perry B W,Miller P,et al.A comparison of methods for the statistical analysis of spatial point patterns in plant ecology[J].Plant Ecol,2006,187:59 -82.
[24]侯向阳,韩进轩.长白山红松林主要树种空间格局的模拟分析[J].植物生态学报,1997,21(3):242-249.
[25]Wiegand T,Moloney K A.Rings,circles,and null- models for point pattern analysis in ecology[J].Oikos,2004,104:209 -229.
[26]苏薇,岳永杰,余新晓.油松天然林群落结构及种群空间分布格局[J].东北林业大学学报,2009,37(3):18-20.
[27]Nakashizuka T.Species coexistence in temperate,mixed deciduous forests[J].Trends Ecol Evol,2001,16:205 - 210.
[28]Dietrich S,Helga S.Fractals,random shapes and point fields:methodsofgeometricalstatistics[M]. New York:John Wiley&Sons,1994.
[29]赵士洞,郝占庆,陶大立.长白山北坡阔叶红松林及其次生白桦林高等植物物种多样性比较[J].应用生态学报,1994,5(1):16-23.
[30]缪宁,刘世荣,史作民,等.川西亚高山红桦—岷江冷杉林优势种群的空间格局分析[J].应用生态学报,2009,20(6):1263-1270.
[31]闫海冰,韩有志,杨秀清,等.华北山地典型天然次生林群落的树种空间分布格局及其关联性[J].生态学报,2010,30(9):2311-2321.
[32]鲁长虎.动物与红松天然更新关系的研究综述[J].生态学杂志,2003,22(1):49-53.
[33]郝占庆,陶大立,赵大昌,等.红松天然更新对动物的依赖性:一个排除动物影响的球果发芽实验[J].生物多样性,1995,8(3):131-133.
[34]徐振邦,戴洪才,李昕.主要伴生树种树叶对红松生长的影响[J].林业科学,1992,28(4):357-361.
[35]刘振国,李镇清.植物群落中物种小尺度空间结构研究[J].植物生态学报,2005,29(6):1020-1028.
