物种分布模型理论研究进展
2013-09-15李国庆刘长成刘玉国张新时
李国庆,刘长成,刘玉国,4,杨 军,5,张新时,郭 柯,*
(1.西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室,杨凌 712100;2.中国科学院植物研究所植被与环境变化国家重点实验室,北京 100093;3.中国科学院水利部水土保持研究所,杨凌 712100;4.中国林业科学研究院荒漠化研究所,北京 100091;5.北京电子科技职业学院生物工程学院,北京 100029
物种分布模型(SDMs)主要是利用物种的分布数据(主要是出现数据)与环境数据,依据特定的算法估计物种的生态位,并投影到景观中,以概率的形式反映物种对生境的偏好程度,结果可以解释为物种出现的概率、生境适宜度或物种丰富度等。它在环境科学的研究、自然资源的管理和生物多样性的保护方面具有重要的应用价值,这些应用包括生物多样性评估、自然保护区的设计、生态恢复中物种的选择、物种迁地保护生境的筛选、环境风险评估、入侵物种的管理、群落和生态系统分布的模拟、全球环境变化对物种和生态系统影响等方面[1-3]。
SDMs的研究起源于早期植物群落与环境梯度关系的研究[4],特别是物种对环境因子响应曲线的研究[5-6]。在20世纪70年代,Nix等最早使用SDMs预测物种的空间分布[7],但这一时期研究SDMs的主要目的还是理解物种分布与环境的关系;在20世纪80年代,由于计算机技术和统计科学的发展,使得SDMs逐渐转向以预测为目的的研究;20世纪90年代以后,GIS技术的快速发展及数字地球表面高程模型、气候的插值数据、海洋和陆地表面遥感数据获得越来越容易,大大加强了SDMs的应用能力,大量的物种分布模型及软件涌现出来(表 1)[8-18]。
中国学者对SDMs的研究起步较晚,20个世纪后期的科学家们主要致力于研究植被类型或物种与气候的定量关系[19-21]。进入21世纪后,学者们的研究目标开始转移至物种潜在分布区预测上[22-27]及气候变化对物种潜在分布区的影响[28-31]。这些研究主要集中在SDMs的应用领域,而对SDMs的理论问题探讨较少[32-34]。本文总结近年来国内外关于SDMs理论的最新进展、存在的理论问题及未来研究的发展方向,为读者更有效的利用和发展SDMs提供参考。

表1 免费使用的物种分布模型软件包Table 1 Free software for modeling species distribution
1 物种分布模型的理论基础
生态位理论在SDMs的研究中始终占据主导地位[35-37]。物种生态位概念具有不同的解释,Grinnell[38]利用限制性环境因子定义的生态位,称为空间生态位;Elton[39]定义的物种生态位是物种在群落中的功能地位,即功能生态位;Hutchinson[40]定义的生态位为物种在环境多维空间中的超体积,在这种环境条件下物种能够生存且繁殖,称为超体积生态位。Hutchinson进一步区分了基础生态位和现实生态位:基础生态位是在没有物种竞争情况下的物种能够生存的环境条件;现实生态位包括了物种之间的相互作用,在这种条件下物种能够生存且繁殖。SDMs与Hutchinson的超体积生态位关系密切,SDMs强调物种的生态需求,特别是重要的非生物因子控制物种的分布,因此在SDMs的研究中通常使用多个环境变量来预测物种的生态位,从而模拟物种的分布区。
关于SDMs是模拟物种的基础生态位还是模拟物种的现实生态位,在过去的研究中受到了很多的争论[41]。Stockwell and Peters[12]认为SDMs模拟的是物种的基础生态位,而不是现实生态位,因为生物的相互作用没有被考虑在模型中;然而Austin[35]和Guisan和Thuiller[2]认为用于建立模型的物种出现数据已经包括了的物种相互作用信息,因此模拟出来的生态位应该是物种的现实生态位;Soberón and Peterson[36]认为在用于建立SDMs的物种出现数据可能包括了来自库种群,这些库种群所在的环境条件不能满足种群延续的需要,因此在这种情况下模型模拟出的生态位既不属于基础生态位也不属于现实生态位;Hizel和Le Lay[37]注意到物种之间的相互作用主要发生在很小距离上的,扩散限制和小尺度的环境异质性允许竞争劣势种在没有竞争者的地方回避负的相互作用,认为实际上区分基础生态位和现实生态位是没有必要的,特别是在尺度较大或精度较粗糙的环境因子条件下,如气候等。因此,SDMs模拟的生态位与物种的生态位的关系是不确定的,还需要深入的研究。
SDMs虽然和生态位理论有重大的关系,但源-库理论和集合种群理论在SDMs的研究中也有非常重要的作用。源-库理论认为可以把物种的分布生境区分成源生境和库生境[42-44],在源生境中种群的维持依靠出生率大于死亡率,而库生境中死亡率大于出生率,种群的维持来自于源生境中物种的扩撒;集合种群理论认为一个物种可能由于局部灭绝而从适合生境的斑块中消失,或者是扩散的限制而未能到达适合的生境[45]。按照这两个理论的认识,物种可能分布在并不适合其生长的环境中(存在于库生境中),或者从适合生境中缺失(局部灭绝)。这个两个理论对于使用SDMs来说是非常重要的,因为大多数的SDMs都是根据观察到的物种出现数据推测物种的环境条件,因此不能简单的把物种出现数据的生境等同于物种适合生境,模型预测的生境适合区域也不一定有物种的存在。
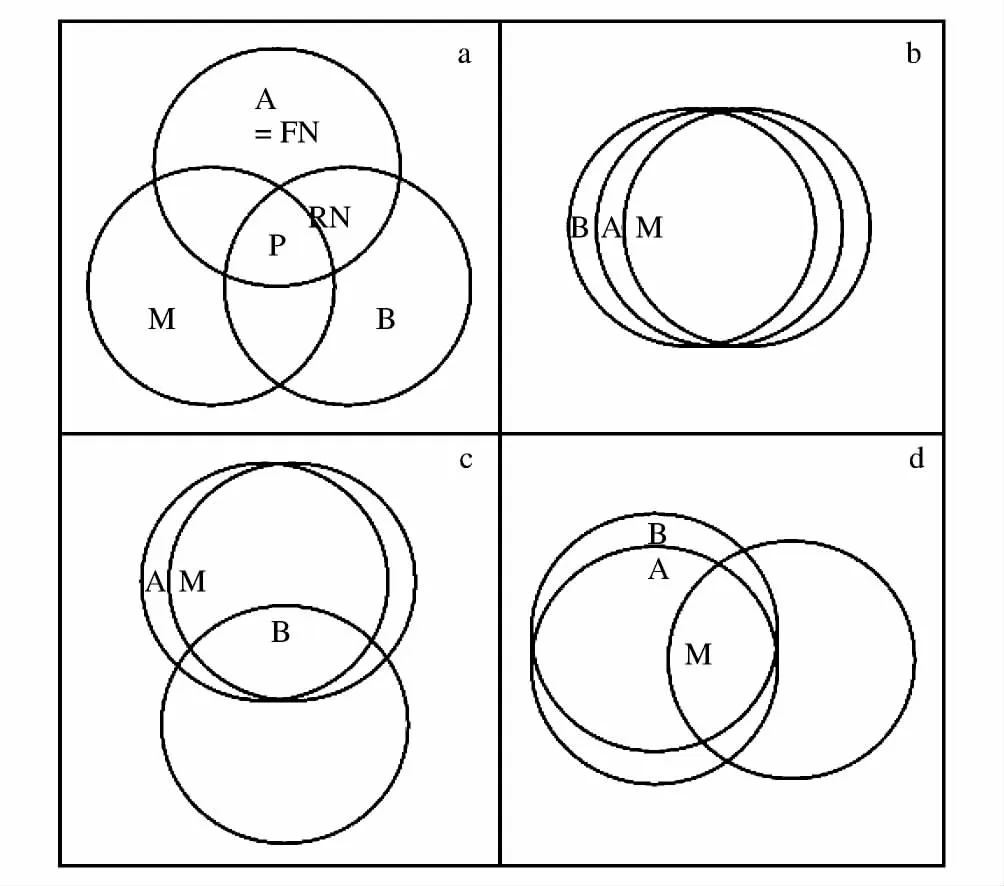
图1 用于解释物种分布模型与物种分布区关系的静态的、非机理的概念化模型(修改Soberón and Peterson,2005)Fig.1 The static, no mechanistic conceptual model for explaining the relationship between species distribution model and species distribution area(modified from Soberón and Peterson,2005)(a)物种的基础生态位、物种之间相互作用和物种的扩散能力的示意图;(b)A、B与M都重合;(c)A仅与M重合,但不与B重合;(d)A仅与B重合,但不与M重合.其中A代表基础生态位(FN)区域,B代表物种之间的相互作用区域,M代表物种扩散的区域,RN代表现实生态位区域,P表示物种实际分布区
2 物种分布模型与物种分布区的关系
物种分布区是物种生态与进化历史的复杂表达形式,在不同的时空尺度上受到多种因素、不同强度的控制[46-47]。Soberón等认为决定物种分布区共有四种因素[36,48-49]:(1)环境因素,包括气候、地形、土壤、生理环境等通过直接或间接的影响物种的生理生态功能,进而限制物种的分布;(2)生物因素,通过与其它物种之间的相互作用(包括互惠、种子扩散、授粉、竞争、捕食、疾病等),导致物种的适应能力变化,从而影响物种的分布区;(3)物种的扩散能力,该因素受到地形障碍而无法到达适合生境,或者能够到达其不适合的生境,在不适合生境中的种群能够生存但不能繁殖;(4)物种适应新环境的进化能力,这种因素的作用通常被认为是很小的,但在物种超出了其可能的分布区的情况下,该种因素可能是非常重要的。
在上述的4种因素中,由于物种适应新环境的进化能力在短时间内很少发生。因此,Soberón等发展了一个静态的、非机理的概念化模型(图1a)来解释其它的三种因素(环境因素A、生物因素B、物种的扩散能力M)对物种分布区的影响[36,49]。通过该模型可以推演出3种情况来解释物种分布模型与物种分布区的关系:(1)当图1中的3个区域(ABM)完全重合时 (图1b)。在这种情况下基础生态位等于现实生态位,因为在这种情况下没有竞争的限制也没有扩散的限制,SDMs估计物种分布区的准确程度完全取决于模型算法的本身和物种在环境A中的取样比例,在取样合理的情况下,SDMs能够准确的模拟物种的分布区;(2)当A仅与M重叠,而与B不重叠时(图1c),即表示物种扩散能力较强或研究区域不存在迁移障碍,物种之间存在强烈的相互作用,物种占有的现实生态位小于基础生态位。当SDMs使用非生物的环境因素来预测物种的分布区时,由于在模型中缺少了物种的相互作用的成分,在这种情况下将高估物种的分布区;(3)当A仅与B重叠,而与M不重叠时(图2d),在这种情况下,物种之间的相互作用较弱,对物种分布模型预测的准确度影响不大。但是,这时由于物种扩散能力较弱或在研究区域存在迁移障碍,物种分布可能呈现出岛屿化状态,其占有的生态位远小于物种的基础生态位和现实生态位,因此,当使用SDMs结合实际出现的物种分布数据模拟物种的分布区将大大低估物种的分布区。
从上述的分析得知,只有在3组影响因素的地理分布完全重合的情况下,物种分布模型能够正确的估计出物种的潜在分布区;在A仅与M重叠的情况下,物种分布模型将高估物种的分布区;在A仅与B重叠的情况下,物种分布模型将低估物种的分布区。从目前的资料来看,这3组影响因素的重叠程度是个经验问题[36],一般说来,如果研究区域面积较小或者研究区域具有同质的生物地理历史过程,生物意义上M应该接近于A;如果区域物种之间的相互作用较弱,A与B期望的重叠度较大。
3 物种分布模型存在的问题与未来的研究方向
3.1 加强模型背后的理论框架的研究
SDMs与生态位有密切的关系,但到目前为止,SDMs模拟的生态位是物种的基础生态位还是现实生态位仍然存在争议[2,12,36-37]。争议的焦点主要是物种的相互作用是否已经被纳入SDMs中?或者物种相互作用对SDMs模拟物种分布区的准确度是否有影响?目前虽然有人认为物种出现数据中已经包括了生物相互作用的信息[2,35],但在SDMs模拟的过程中能否得到充分反映呢?是不清楚的。物种的相互作用在群落生态学的零模型假设中备受争议,中性模型认为物种具有相同的生态需求并且相互之间没有竞争,在这种情况下是足以描述物种的分布格局;生态位理论认为生态位相同的物种因竞争相同的资源而不能稳定共存[50-51]。目前的生态学家一致认为这两种过程共同作用于物种的分布。因此,在未来SDMs的研究中应该考虑纳入物种相互作用过程。在整合生物相互作用的过程中,应该充分重视尺度对生物的相互作用的影响[52-53],因为有些生态过程可能发生在较大或较粗的尺度上,而不发生在较小或较细的尺度上,例如在较大的尺度上,竞争或互利应该对物种的分布具有较小的影响[37,52-53],然而却对局地物种丰富度产生影响。除了竞争和互利之外,捕食、寄生、病原菌、互惠等作用对物种的分布影响也具有重要作用。
用于SDMs研究的物种分布数据,一般情况下是从实际的野外调查中获得的或者从标本、文献记载获得的[54]。搜集的物种出现数据可能已经包括了来自库生境中的种群,这种情况下SDMs模拟的生态位既不是基础生态位,也不是现实生态位[43,55]。物种分布数据是来自于源生境还是库生境的判断标准是物种在该地区是否能够长期生存且顺利繁殖。因此,在未来的SDMs研究中应该更多的考虑物种的生殖的成功率,这就需要一个更综合的理论框架来整合生态位理论和源-库理论。
3.2 在模型中整合种群动态过程与物种迁移过程
SDMs模型假定物种与环境之间达到一种平衡的状态,这是使用SDMs预测的核心假设[35,56],但物种分布的实际情况是否如此?或者在环境变化的情况下,物种重新达到平衡状态需要多长时间?这就需要我们在SDMs模型中整合种群的动态过程来解决上述问题。除此之外,在模型中整合种群动态过程不仅可以帮助我们理解SDMs模型预测产生的Commission和Omission错误的生物学原因,而且能够为集合种群的研究提供有力的技术支持[2]。
SDMs能够模拟出物种在气候变化情景下的适合生境,却没有考虑物种是否能够赶上气候变化的速度[57-59]。因此有时模拟的结果与实际的情况相差较大,可能是由于物种本身的迁移能力较弱,或者景观中存在迁移障碍导致物种不能到达其适合的生境[60]。因此在SDMs模型中加入物种的迁移过程就能够克服上述问题,能更真实的模拟物种随环境或气候变化的动态过程[61]。在未来的研究中,两个方面的研究不应该被忽略,一是模拟随机的长距离扩散事件对物种分布的影响和二是迁移障碍对物种扩散的影响。
3.3 加强模型与生态位进化理论相互关系的研究
物种分布数据一般都是从物种整个分布区获得的,它们全面的覆盖了整个物种分布区的环境条件。但这些工作忽略了物种出现数据是否代表一个单一的进化整体或者一个集合的进化谱系,物种是否能够随着年龄、进化的独立性、遗传的独特性而改变[62]。进而引出两个问题:物种下分类水平的亚种群内部的生态位是否出现分化了呢?或者物种的基础生态位是否已经进化了呢?研究表明广泛分布的种群在空间连续的区域能够表现出持续的形态特征差异,这种变异与所研究的基因位点的变异或局地适应不一致,表明有受到选择作用的基因位点存在[63-64];也有研究表明独立的分类群的基础生态位能够数千年保持恒定[65],或能够在短短的数代内大幅的发生改变[66]。相关物种的生态位漂移和保守的证据是模糊不清的或存在争论的[67-68]。而物种生态位是否保守对SDMs模型的成功预测至关重要,特别是评估气候变化对物种潜在分布的影响,因此以后应该加强物种生态位稳定性和进化研究。
3.4 从更高理论层次加强功能群和群落结构的模拟
从理论上说,预测单个物种的潜在分布区可以分析更高水平的生态复杂度,目前这类的SDMs研究报道相对较少,这里的一个主要困难是物种聚合的规则,特别是预测未来气候变化情况下物种的聚合规则[69-70]。目前可以归纳出如下3种方法:1)先聚合物种,再预测群落分布;2)先预测物种分布,再聚合群落分布;3)聚合和预测同时进行。第一种方法是在知道物种的聚合规则的前提下进行预测的,如实地调查群落的组合物种;第二种情况是在未知物种聚合规则情况下,来推测群落的物种聚合规则的,不考虑物种之间的相互作用;第三种情况是根据物种共同出现的情况进行预测的,包括了物种之间的相互作用[71-72]。这3个群落聚合规则与群落生态学中的有机体论和个体论学说不谋而合,先聚合再预测的策略符合群落有机体论学说;先预测再聚合符合群落个体论学说;聚合和预测同时进行符合群落有机体论-个体论综合的学说。探索和发展这3种物种聚合规则在功能群和群落结构模拟中的应用前景,必将是未来SDMs研究中的热点问题[73-75]。
总之,SDMs应该更多的联系生态学中的理论,如生态位理论、种群动态理论、源-库理论、集合种群理论、进化理论及群落构建理论等。相信通过科学家的努力,未来的SDMs应该能够动态的描述多物种的潜在分布区、更能深入到生态学的理论研究中。
[1] Franklin J.Predictive vegetation mapping:geographic modelling of biospatial patterns in relation to environmental gradients.Progress in Physical Geography,1995,19(4):474-499.
[2] Guisan A,Thuiller W.Predicting species distribution:offering more than simple habitat models.Ecology Letters,2005,8(9):993-1009.
[3] Elith J,Leathwick J R.Species distribution models:ecological explanation and prediction across space and time.Annual Review of Ecology,Evolution,and Systematics,2009,40:667-697.
[4] Grinnell J.The origin and distribution of the chestnut-backed chickadee.The Auk,1904,21(3):364-365.
[5] Whittaker R J.Vegetation of the Great smoky mountains.Ecological Monography,1956,26(1):1-80.
[6] MacArthur R H.Population ecology of some warblers of northeastern coniferous forests.Ecology,1958,39(4):599-619.
[7] Nix H,MacMahon J,Mackenzie D.Potential areas of production and the future pigeon pea and other grain legumes in Australia//Wallis E S,Whiteman P C,eds.The potential for pigeon pea in Australia.Proceedings of Pigeon Pea(Cajanus cajan(L.)Millsp.)Field Day.Queensland,Australia:University of Queensland,1977:1-12.
[8] Aspinall R,Veitch N.Habitat mapping from satellite imagery and wildlife survey data using a Bayesian modeling procedure in GIS.Photogrammetric Engineering and Remote Sensing,1993,59:537-543.
[9] Hizel A H,Hausser J,Chessel D,Perrin N.Ecological niche factor analysis:how to compute habitat suitability maps without absence data?Ecology,2002,83(7):2027-2036.
[10] Thuiller W.BIOMOD-optimizing predictions of species distributions and projecting potential future shifts under global change.Global Change Biology,2003,9(10):1353-1362.
[11] Hijmans R J,Guarino L,Cruz M,Rojas E.Computer tools for spatial analysis of plant genetic resources data:1.DIVA-GIS.Plant Genetic Resources Newsletter,2001,127:15-19.
[12] Stockwell D,Peters D.The GARP modelling system:problems and solutions to automated spatial prediction.International Journal of Geographical Information Science,1999,13(2):143-158.
[13] Lehmann A,Overton J,Leathwick JR.GRASP:Generalized regression analysis and spatial prediction.Ecological Modelling,2002,157(2/3):189-207.
[14] McCune B.Nonparametric habitat models with automatic interactions.Journal of Vegetation Science,2006,17(6):819-830.
[15] Farber O,Kadmon R.Assessment of alternative approaches for bioclimatic modeling with special emphasis on the Mahalanobis distance.Ecological Modelling,2003,160(1/2):115-130.
[16] Phillips SJ,Anderson R P,Schapire R E.Maximum entropy modeling of species geographic distributions.Ecological Modelling,2006,190(3/4):231-259.
[17] Guo Q H,Liu Y.ModEco:an integrated software package for ecological niche modeling.Ecography,2010,33(4):637-642.
[18] Rangel T F L V B,Diniz-Filho JA F,Bini L M.Towards an integrated computational tool for spatial analysis in macroecology and biogeography.Global Ecology and Biogeography,2006,15(4):321-327.
[19] Hong B G,Li S Z.The preliminary study of the correlations between the distribution of main everygreen broad-leaf tree species in Jiangsu and climats.Acta Ecologica Sinica,1981,1(2):105-111.
[20] Wang X R,Fan J H,Wang X S.Distribution of main tree species and its relation to water and heat conditions in shelter-forest districts of“San Bei”.Chinese Journal of Ecology,1986,5(1):13-17.
[21] Zhang X S.A vegetation-climate classification system for global change studies in China.Quaternary Sciences,1993,13(2):157-169.
[22] Jiang X,Ni J.Species-climate relationships of 10 desert plant species and their estimated potential distribution range in the arid lands of northwestern China.Acta Phytoecologica Sinica,2005,29(1):98-107.
[23] Cao M C,Zhou G S,Wang E S.Application and comparison of generalized models and classification and regression tree in simulating tree species distribution.Acta Ecologica Sinica,2005,25(8):2031-2040.
[24] Li F,Zhou G S,Cao M C.Responses of Larix gmelinii geographical distribution to future climate change:a simulation study.Chinese Journal of Applied Ecology,2006,17(12):2255-2260.
[25] Zhang Z D,Zang R G.Predicting potential distributions of dominaint woody plant keystone species in a natural tropical forest landscape of Bawangling Hainan Island South China.Journal of Plant Ecology(Chinese Version),2007,31(6):1079-1091.
[26] Xu X,Yang Y,Wang L.Geographic distribution and potential distribution estimation of Pseudotaxus chienii.Chinese Journal of Plant Ecology,2008,32(5):1134-1145.
[27] Peng SZ,Zhao C Y,Xu Z L,Wang C,Liu Y Y.Potential distribution of Qinghai spruce and assessment of its growth status in the upper reaches of the heihe river in the Qilian Mountains of China.Chinese Journal of Plant Ecology,2011,35(6):605-614.
[28] Xu Z L,Zhao C Y,Feng Z D.A study of the impact of climate change on the potential distribution of Qinghai spruce(Picea crassifolia)in Qilian Mountains.Acta Ecologica Sinica,2009,29(5):278-285.
[29] Wang J,Ni J.Modelling the distribution of five Caragana species in temperate northern China.Chinese Journal of Plant Ecology,2009,33(1):12-24.
[30] Wu J G,Lü J J.Potential effects of climate change on the distribution of dove trees(Davidia involucrata Baill)in China.Research of Environmental Sciences,2009,22(12):1371-1381.
[31] Zhang L,Liu SR,Sun P S,Wang T L.Comparative evaluation of multiple models of the effects of climate change on the potential distribution of Pinus massoniana.Chinese Journal of Plant Ecology,2011,35(11):1091-1105.
[32] Zuo W Y,Lao N,Geng Y Y,Ma K P.Predicting species's potential distribution-SVM compared with GARP.Journal of Plant Ecology(Chinese Version),2007,31(4):711-719.
[33] Wang Y S,Xie B Y,Wan F H,Xiao Q M,Dai L Y.Application of ROC curve analysis in evaluating the performance of alien species'potential distribution models.Biodiversity Science,2007,15(4):365-372.
[34] Shao H,Tian JQ,Guo K,Sun JX.Effects of sample size and species traits on performance of BIOCLIM in predicting geographical distribution of tree species-a case study with 12 deciduous Quercus species indigenous to China.Chinese Journal of Plant Ecology,2009,33(5):870-877.
[35] Austin M P.Spatial prediction of species distribution:an interface between ecological theory and statistical modeling.Ecological Modelling,2002,157(2/3):101-118.
[36] Soberón J,Peterson A T.Interpretation of models of fundamental ecological niches and species's distribtional areas.Biodiversity Informatics,2005,2:1-10.
[37] Hizel A H,Le Lay G.Habitat suitability modeling and niche theory.Journal of Applied Ecology,2008,45(5):1372-1381.
[38] Grinnell J.The niche-relationships of the California thrasher.The Auk,1917,34(4):427-433.
[39] Elton C.Animal Ecology.Chicago:University of Chicago Press,1927.
[40] Hutchinson M.Methods for generation of weather sequences//BuntingA H ed.Agricultural environments:characterisation,classification and mapping.Wallingford:CAB,International,1987:149-157.
[41] Sillero N.What does ecological modelling model?A proposed classification of ecological niche models based on their underlying methods.Ecological Modelling,2011,222(8):1343-1346.
[42] Pulliam H.Sources,sinks,and population regulation.American Naturalist,1988,132(5):652-661.
[43] Pulliam H.On the relationship between niche and distribution.Ecology Letters,2000,3(4):349-361.
[44] Dias P C.Sources and sinks in population biology.Trends in Ecology and Evolution,1996,11(8):326-330.
[45] Hanski I.Metapopulation Ecology.Oxford,UK:Oxford University Press,1999.
[46] Brown JH.Macroecology.Chicago:University of Chicago Press,1995.
[47] Franklin J.Mapping Species Distributions:Spatial Inference and Prediction.Cambridge,UK:Cambridge University Press,2009.
[48] Soberón J M.Niche and area of distribution modeling:a population ecology perspective.Ecography,2010,33(1):1-9.
[49] Soberón JM,Nakamura M.Niches and distributional areas:concepts,methods and assumptions.Proceedings of the National Academy of Sciences of the United States of America,2009,106(17):19644-19650.
[50] Hubbell S.Neutral theory and the evolution of ecological equivalence.Ecology,2006,87(6):1387-1398.
[51] Bell G.Neutral macroecology.Science,2001,293(5539):2413-2418.
[52] Kriticos D J,Leriche A.The effects of climate data precision on fitting and projecting species niche models.Ecography,2010,33(1):115-127.
[53] Hui C,Veldtman R,McGeoch M A.Measures perceptions and scaling patterns of aggregated species distributions.Ecography,2010,33(1):95-102.
[54] Graham C,Ferrier S,Huettman F,Moritz C,Peterson A T.New developments in museum-based informatics and applications in biodiversity analysis.Trends in Ecology and Evolution,2004,19(9):497-503.
[55] Barve N,Barve V,Jimenez-Valverde A,Lira-Noriega A,Maher SP,Peterson A T,Soberon J,Villalobos F.The crucial role of the accessible area in ecological niche modeling and species distribution modeling.Ecological Modelling,2011,222(11):1810-1819.
[56] Jeschke J,Strayer D.Usefulness of bioclimatic models for studying climate change and invasive species.Annals of the New York Academy of Sciences,2008,1134(1):1-24.
[57] Davis A,Jenkinson L,Lawton J,Shorrocks B,Wood S.Making mistakes when predicting shifts in species range in response to global warming.Nature,1998,391(6669):783-786.
[58] Malcolm JR,Markham A,Neilson R P,Garaci M.Estimated migration rates under scenarios of global climate change.Journal of Biogeography,2002,29(7):835-849.
[59] Engler R,Randin C F,Vittoz P,Czaka T,Beniston M,Zimmermann N E,Guisan A.Predicting future distributions of mountain plants under climate change:does dispersal capacity matter.Ecography,2009,32(1):34-45.
[60] Iverson L,Schwartz M W,Prasad A M.Potential colonization of newly available tree-species habitat under climate change-an analysis for five eastern USspecies.Landscape Ecology,2004,19(7):787-799.
[61] Engler R,Hordijk W,Guisan A.The MIGCLIM R package-seamless integration of dispersal constraints into projections of species distribution models.Ecography,2012,35(10):872-878.
[62] Zimmermann N,Edwards T,Graham C,Pearman P,Svenning J.New trends in species distribution modelling.Ecography,2010,33(6):985-989.
[63] Maron J L,Vilá M,Bommarco R,Elmendorf S,Beardsley P.Rapid evolution of an invasive plant.Ecological Monographs,2004,74(2):261-280.
[64] Wright J W,Davies K F,Lau J A,McCall A C,McKay J K.Experimental verification of ecological niche modeling in a heterogeneous environment.Ecology,2006,87(10):2433-2439.
[65] Peterson A,Soberon J,Sanchez-Cordero V.Conservatism of ecological niches in evolutionary time.Science,1999,285(5431):1265-1267.
[66] Clements D R,Ditommaso A.Predicting weed invasion in Canada under climate change:evaluating evolutionary potential.Canadian Journal of Plant Science,2012,92(6):1013-1020.
[67] Petitpierre B,Kueffer C,Broennimann O,Randin C,Daehler C,Guisan A.Climatic niche shifts are rare among terrestrial plant invaders.Science,2012,335(6074):1344-1348.
[68] Webber B L,Le Maitre D C,Kriticos D J.Comment on“climatic niche shift are rare among terrestrial plant invaders”.Science,2012,338(6104):193-193.
[69] Ferrier S,Guisan A.Spatial modelling of biodiversity at the community level.Journal of Applied Ecology,2006,43(3):393-404.
[70] Guisan A,Rahbek C.SESAM-a new framework integrating macroecological and species distribution models for predicting spatio-temporal patterns of species assemblages.Journal of Biogeography,2011,38(8):1433-1444.
[71] Baselga A,Araújo M B.Do community-level models describe community variation effectively.Journal of Biogeography,2010,37(10):1842-1850.
[72] Götznberger L,de Bello F,Bråthen K A,Davison J,Dubuis A,Guisan A,LepšJ,Lindborg R,Moora M,Partel M,Pellissier L,Pottier J,Vittoz P,Zobel K,Zobel M.Ecological assembly rules in plant communities-approaches patterns and prospects.Biological Reviews,2012,87(1):111-127.
[73] Cabral J S,Kreft H.Linking ecological niche community ecology and biogeography:insights from a mechanistic niche model.Journal of Biogeography,2012,39(12):2212-2224.
[74] Kissling W D,Dormann CF,Groeneveld J,Hickler T,Kühn I,Mclnerny GJ,Montoya JM,Romermann C,Römermann C,Schiffers K,Schurr F M,Singer Ar,Svenning J C,Zimmermann N E,O'Hara R B.Towards novel approaches to modelling biotic interactions in multispecies assemblages at large spatial extents.Journal of Biogeography,2012,39(12):2163-2178.
[75] Ohmann J L,Gregory M J,Henderson E B,Roberts H M.Mapping gradients of community composition with nearest-neighbour imputation:extending plot data for landscape analysis.Journal of Vegetation Science,2011,22(4):660-676.
参考文献:
[19] 洪必恭,李绍珠.江苏主要常绿阔叶树种的分布与热量关系的初步研究.生态学报,1981,1(2):105-111.
[20] 王效瑞,范建华,汪祥森.“三北”防护林地区主要树种的分布与水、热条件的关系.生态学杂志,1986,5(1):13-17.
[21] 张新时.研究全球变化的植被-气候分类系统.第四纪研究,1993,13(2):157-169.
[22] 蒋霞,倪健.西北干旱区10种荒漠植物地理分布与大气候的关系及其可能潜在分布区的估测.植物生态学报,2005,29(1):98-107.
[23] 曹铭昌,周广胜,翁恩生.广义模型及分类回归树在物种分布模拟中的应用与比较.生态学报,2005,25(8):2031-2040.
[24] 李峰,周广胜,曹铭昌.兴安落叶松地理分布对气候变化响应的模拟.应用生态学报,2006,17(12):2255-2260.
[25] 张志东,臧润国.海南岛霸王岭热带天然林景观中主要木本植物关键种的潜在分布.植物生态学报,2007,31(6):1079-1091.
[26] 徐晓婷,杨永,王利松.白豆杉的地理分布及潜在分布区估计.植物生态学报,2008,32(5):1134-1145.
[27] 彭守璋,赵传燕,许仲林,王超,柳逸月.黑河上游祁连山区青海云杉生长状况及其潜在分布区的模拟.植物生态学报,2011,35(6):605-614.
[29] 王娟,倪健.中国北方温带地区5种锦鸡儿植物的分布模拟.植物生态学报,2009,33(1):12-24.
[30] 吴建国,吕佳佳.气候变化对珙桐分布的潜在影响.环境科学研究,2009,22(12):1371-1381.
[31] 张雷,刘世荣,孙鹏森,王同立.气候变化对马尾松潜在分布影响预估的多模型比较.植物生态学报,2011,35(11):1091-1105.
[32] 左闻韵,劳逆,耿玉英,马克平.预测物种潜在分布区——比较SVM与GARP.植物生态学报,2007,31(4):711-719.
[33] 王运生,谢丙炎,万方浩,肖启明,戴良英.ROC曲线分析在评价入侵物种分布模型中的应用.生物多样性,2007,15(4):365-372.
[34] 邵慧,田佳倩,郭柯,孙建新.样本容量和物种特征对BIOCLIM模型模拟物种分布准确度的影响——以12个中国特有落叶栎树种为例.植物生态学报,2009,33(5):870-877.
