不同土壤水分条件下杨树人工林水分利用效率对环境因子的响应
2013-09-15张志强方显瑞查同刚王小平陈俊崎陈吉泉
周 洁,张志强,*,孙 阁,方显瑞,查同刚,张 燕,王小平,陈俊崎,陈吉泉
(1.北京林业大学水土保持与荒漠化防治教育部重点实验室,北京100083;2.Eastern Forest Environmental Threat Assessment Center,USDA Forest Service,Raleigh,NC 27606,U.S.A;3.北京市林业碳汇工作办公室,北京100085;4.Landscape Ecology&Ecosystem Science,University of Toledo,Toledo,OH 43606-3390,U.S.A)
杨树是我国人工林造林面积最大的树种[1],它被广泛用于用材林、城市绿化以及碳汇造林等诸多方面。杨树生长快,易于繁殖,拥有较高的生产力和生物量,但同时也通过蒸腾和截留作用消耗着大量的水分,是耗水相对较高的树种之一。水是限制陆地生物活动的重要因子之一[2],尤其在干旱半干旱地区水资源对陆地生物影响更为突出[3]。生态系统水分利用效率(WUE)作为表征生态系统消耗单位水量所获得生产力的指标[4],能够更好地量化生态系统碳固定和水分消耗的关系。因此,正确分析水分对杨树人工林WUE的影响对深入了解杨树人工林生产能力和资源损耗的关系,以及对实施植树造林政策、进行森林管理有着重要意义。
随着环境问题的日益突出,气候变化对生态系统碳水循环及其耦合关系的影响得到广泛关注。全球通量网(FLUXNET)的建立为生态系统碳水通量研究提供了大量信息[5-6]。研究者对不同生态系统WUE的变化趋势及影响因子进行了分析,结果显示,影响WUE变化的因素大致分为三类,即植被因素、环境因素和区域差异因素[7-12]。近年来,国外对杨树人工林碳水通量及其关系进行了相关研究。Migliavacca等运用涡度相关法对杨树人工林WUE进行了季节和年际变化的分析,认为随着干旱的到来,生态系统蒸发散(ET)变化不大,而总生态系统生产力(GEP)却有较大下降,因此WUE减小[13]。然而,国内对杨树人工林生态系统尺度WUE的研究还较为有限,并且多数研究将杨树生态系统碳水通量分开研究[14-15],并未在生态系统尺度分析两者的耦合关系。
本文运用涡度相关技术,于2007年对位于北京市大兴区永定河沿河沙地杨树人工林进行了连续动态测定,通过研究GEP和ET随相对土壤含水量(REW)的变化趋势,探讨杨树人工林生态系统不同土壤水分条件下碳水通量的环境响应。本文主要研究目的在于:(1)了解不同水分条件下,生态系统碳-水间的耦合关系;(2)分析水分亏缺对总生态系统生产力、蒸发散及两者关系的影响;(3)探讨碳水通量与环境因子、植物自身气孔行为的关系,以期认识各因子在不同水分条件下对碳-水通量及其关系的调控作用,旨在为提高水分利用效率的杨树集约栽培和经营管理提供理论依据。
1 材料及方法
1.1 研究区概况
试验地位于北京市南郊大兴区榆垡镇大兴林场,属暖温带亚湿润气候区。年平均气温为11.6℃,极端最低气温-27.4℃,极端最高温度40.6℃;多年平均降雨为556mm(1990—2009年),其中7—9月份降雨占全年降雨总量的60%—70%(大兴气象站,116°15'07″E,39°31'50″N,1956—2000年观测数据)。试验区为海河水系永定河洪积区,地势平坦,平均海拔30m,坡度<5°。土壤为冲积性沙壤土,通透性好,保肥蓄水能力差,土壤pH值为8.25—8.39,容重1.43—1.47g/cm3。地下水位年平均降幅达3.9%。
试验区主要为集约栽培的欧美107杨树人工纯林,75%为2002年种植,其余为1998年、2001年和2003年种植,株行距为2 m×2 m,林分均匀整齐。2007年林木平均胸径为12.2cm,平均树高为13m,叶面积指数为2.66m2/m2。林下植物种类较少,多为草本植物,少有灌木,优势种为灰绿藜(Chenopodium glaucum),伴生有紫花苜蓿(Medicago sativa),黄香草木樨(Melilotus officinalis),猪毛菜(Salsola collina),蒺藜(Tribulus terrestris)等。
1.2 试验方法
该试验区下垫面平坦均匀,符合涡度相关法观测要求。利用涡度相关观测系统,在试验站点中心架设一高22.5m的观测塔。主要观测仪器包括净辐射仪(REBS,Seattle,WA)、日照强度计(LI200 X-L,Li-Cor,NE)、光量子传感器(LI190 SB-L,Li-Cor,NE)、H2O/CO2红外气体分析仪(LI-7500,Li-Cor,NE)、三维超声风速仪(CSAT3,CS,USA),以上各仪器安装高度均为18m;气压计(CS105,CS,USA)和翻斗式自动雨量计(TE525-L,CS,USA)安装高度分别为21m和22.5m;空气温湿度传感器(HMP45C probe,CS,USA)在5、10、15、20m高度处各安装一套;土壤温度传感器(TCAV107,CS,USA)和土壤热通量板(HFT3,Seattle,WA)均置于地表以下5、10、20cm处;土壤水分观测仪TDR(CS616,CS,USA)位于地表以下5cm和20cm处。所有气象资料均采用数据采集器(CR5000,CS,USA)每30min自动记录1次。
1.3 数据处理和计算
1.3.1 数据处理及生态系统GEP、ET和WUE的计算方法
由于气象、地形、植被和仪器系统等的影响,涡度相关系统观测获得的数据必须进行质量控制和质量分析。为了简化流程并将大量数据处理程序化,运用EC-processor程序[16]自动实现涡度相关系统数据的剔除、坐标转换、质量控制和质量分析。潜热通量和显热通量数据经过WPL矫正[17],消除水热传输造成的CO2通量变化的密度效应,然后通过三维坐标旋转矫正三维风速,从而使得垂直风向风速的平均值为零,水平风速方向与主导方向一致。
本研究将WUE定义为GEP和ET的比值。用涡度相关技术观测的碳通量为净生态系统碳交换量(NEE),它与净生态系统生产力(NEP)的绝对值相等,但符号相反。即:GEP=-NEE+Re,式中Re为生态系统呼吸总量。在以往的研究中经常使用仅将温度作为生态系统呼吸影响因子的[18]公式,然而,由于土壤呼吸的复杂性,只考虑温度因素不足以正确的描述土壤呼吸对环境的响应[19],本研究采用Law model指数方程[20]来计算生态系统呼吸,同时考虑温度与土壤水分的综合影响。
1.3.2 冠层导度(Gs)的计算方法
ET对环境因子的响应主要受到冠层导度(Gs)的影响,本文根据Penman-Monteith方程计算植被在冠层尺度对水汽的导度[21]:
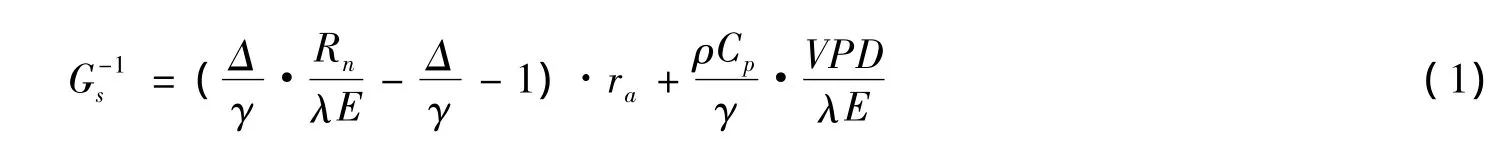
式中,Gs为冠层导度(m/s),Δ为空气饱和水汽压差随温度的变化斜率(kPa/K),γ为干湿球常数(kPa/K),Rn为观测高度的净辐射(W/m2),λE为涡度相关技术观测的潜热通量(W/m2),ra为大气边界层的空气动力学阻抗(s/m),ρ为空气密度(kg/m3),Cp为在恒定压力下的空气热容(kJkg-1K-1),VPD为空气饱和水汽压差(kPa)。其中,ra是根据Monin-Obukhov的相似理论和中性条件下的假设来估算的,式中,Z为风速的观测高度(m),d为零平面位移(m),z0为动量粗糙度(m),z0v为水汽压的粗糙度(m),κ为卡门常数,一般取值为0.4,μ为观测高度的风速(m/s)。本文零平面位移d假定为0.8倍冠层高度,z0为0.1倍冠层高度,z0v为0.01倍冠层高。

1.3.3 相对土壤含水量(REW)
由于土壤质地不同,土壤含水量并不是最好的衡量生态系统水分多寡的标准,相对土壤含水量(REW)成为衡量土壤水分的良好指标。即:REW=(VWC-VWCmin)/(VWCmax-VWCmin),式中VWC为土壤体积含水量(%),VWCmin和VWCmax为研究中最小和最大土壤体积含水量(%)。研究认为,当REW<0.4时,生态系统受到土壤水分胁迫的影响[22-23]。
1.3.4 能量闭合
对系统内能量闭合程度分析是检验EC法观测数据质量的有效手段[24-26],本文采用潜热通量和显热通量之和(LE+Hs)与可提供能量(Rn-G)两者的线性回归方程方法,对2007年该生态系统经质量控制后的30min数据进行能量闭合分析,所得回归直线斜率为0.78(R2=0.84,N=13950,P<0.0001),验证结果说明该站点的涡度相关法观测数据是可靠的[25]。
2 结果与分析
2.1 环境因子季节变化特征
图1为研究区2007年环境因子季节变化特征图。从图中看出,研究区冬季寒冷干燥,夏季高温多雨,各环境因子具有明显的季节变化特征。2007年日平均气温变化范围为-9.7—29.4℃,其中,最高温度出现在7月份,年平均气温为12.9℃,高于多年平均温度11.6℃(1990—2009年),此外,5月份气温比多年同期平均水平高(2±0.5)℃(1990—2009年)。2007年为湿润年,年降雨量为661mm,略高于多年平均降雨量556mm(1990—2000年),主要集中在7—9月份,然而,该研究区6月经历了高温少雨天气,月降雨量为6.4mm,仅为多年平均水平的10%(1990—2009年)。光合有效辐射(PAR)从冬季到夏季逐渐增加,随后逐渐降低。大气饱和水汽压差(VPD)在温度较低的冬季达到极小值,在高温干旱条件下达到极大值。土壤水分(VWC)与降雨量变化较为一致,生长季保持在4.5%—15.0%之间,6月份出现夏季干旱,土壤水分仅为3%。

图1 研究地2007年降雨量、光合有效辐射、大气温度、土壤体积含水量(20cm)和大气饱和水汽压差的季节变化Fig.1 The seasonal variation of precipitation(P),photosynthetically available radiation(PAR),air temperature(Ta),soil volume moisture content(VWC;20cm)and saturated vapor pressure deficit(VPD)at the polar plantation in Daxing site of Beijing in 2007
2.2 土壤水分对水分利用效率的影响
GEP和ET之间存在线性相关性,二者斜率可做为衡量生态系统水分利用效率的标准[27]。基于2007年生长季日尺度数据进行的相关分析发现,GEP和ET在不同土壤水分条件下存在较为显著的线性相关性(REW<0.1:R2=0.86,P<0.001;0.1<REW<0.4:R2=0.51,P<0.001;REW>0.4:R2=0.52,P<0.001),GEP随ET的增加而增大,但两者斜率(WUE)随着土壤水分的增加而逐渐减小(REW<0.1∶2.1;0.1<REW<0.4∶1.7;REW>0.4∶1.5)(图2)。表1可知,REW<0.1,生态系统由于受到了严重的水分限制,GEP和ET维持在较低水平,随着土壤水分的增加,生态系统GEP和ET明显增大,但GEP的增加幅度小于ET,导致WUE减小。0.1<REW<0.4,生态系统GEP与未受土壤水分胁迫条件下GEP相似,但ET却低于水分充足条件,因此WUE表现出增加趋势,说明土壤水分并未被植物有效利用。
相关研究得到相似结果,认为干旱胁迫对蒸散的影响快于光合[28-29]。光合作用除受到气孔的限制外,还受到内部CO2扩散阻力(运送和羧化作用阻力)的影响,因此,在水分胁迫条件下植物的WUE值高[30]。但其他研究却发现,随着夏季干旱的到来,ET值变化不大,而GEP却有较大的下降,因此WUE随水分胁迫有减小趋势[13,31-32]。还有研究认为,随土壤水分含量的减小,ET和GEP同时减小,因此WUE变化不大[33]。Migliavacca认为,造成上述差异的原因主要与地下水位的高低有关[13],即使干旱,只要地下水位和植被根部相近,土壤也能为植被提供足够的水分来补偿不断消耗的水分。在本研究中,沙地具有保水能力差的特性,较差的土壤保水能力导致充足的土壤水分并未被植物有效利用,而通过蒸发散回到大气中,因此造成在土壤水分胁迫条件下,WUE反而高的结果。

图2 2007年生长季(5—10月)不同土壤水分条件下总生态系统生产力和生态系统蒸发散关系图Fig.2 Relationship between gross ecosystem productivity(GEP)and evaportranspiration(ET)at different water conditions in growing seasons of 2007

表1 3种土壤水分条件下生态系统平均总初级生产力、蒸发散和水分利用效率对比Table 1 Comparison of gross ecosystem productivity(GEP),evaportranspiration(ET)and water-use efficiency among different water conditions in the growing season of 2007
2.3 不同土壤水分条件下水分利用效率对环境因子的响应
环境因素是影响生态系统碳水耦合的重要驱动因子。为了进一步分析该生态系统碳固定和水分蒸发散的控制机制,该文分别探讨不同土壤水分条件下碳水通量对环境因子的响应特征。生态系统GEP和ET的变化主要受到气象因子和下垫面条件的影响,气象因子主要包括:气温(Ta)、大气饱和水汽压差(VPD)、光合有效辐射(PAR);下垫面因子为土壤水分(VWC)和土壤温度(Ts)。对生长季半小时数据,采用多元线性逐步回归分析中自后淘汰变量法,得到不同土壤水分条件下,生态系统GEP、ET和WUE与各因子的综合模型,

REW<0.1,生态系统GEP和ET维持在较低水平,各环境因子与其没有显著的相关性(R2<0.2,P>0.1),综合模型显著性较差(P=0.095)。说明在极度干旱条件下,较低的土壤水分限制了植物的生理活动。由方程可以看出,当0.1<REW<0.4时,生态系统GEP和ET分别与VWC呈正相关,生态系统WUE与VWC呈负相关。当生态系统未受到土壤水分胁迫时,生态系统GEP主要受到Ta和PAR的影响,ET主要受到Ta和VPD的影响,而WUE则主要与VPD呈负相关。

图3 0.1<REW<0.4条件下,土壤水分对生态系统总生产力,蒸发散和水分利用效率的影响,误差线为标准差Fig.3 The effects of soil volume water moisture content in 20cm depth(VWC,20cm)on gross ecosystem productivity(GEP),evaportranspiration(ET)and water use efficiency(WUE)at 0.1<REW<0.4 conditions in the growing season of 2007,the error bar represent the SD
0.1<REW<0.4,生态系统GEP和ET主要受到土壤水分影响,而气侯因子对生态系统碳固定和水分蒸发散影响不显著(R2<0.35,P>0.1)。这一结果与宋霞等对亚热带人工林水分利用效率的研究较为一致,研究显示,夏季干旱期,GEP和ET对土壤湿度变化较为敏感,而与净辐射之间没有显著的相关性[34]。虽然GEP和ET随着VWC的增加而增大,但由于ET对土壤水分变化更为敏感,导致ET随VWC增大幅度大于GEP(图3),因此WUE随VWC增大而减小(图3)。土壤水分胁迫对气孔导度造成影响[35],而气孔导度对ET的影响快于GEP[28-29],因此WUE随着VWC的减小而增大。
当REW>0.4时,生态系统GEP和ET主要受到气象因子的影响,各因子通过控制气孔对冠层蒸散以及碳固定产生影响,因此各气象因子与冠层导度的相关性是研究其与生态系统碳水通量及两者关系的关键。如图4所示,冠层导度与气温(Ta)(R2=0.65,P<0.001)和光合有效辐射(PAR)呈显著的正相关关系(R2=0.43,P<0.001)。VPD综合反映了大气中的湿度和温度状况,其与Gs的关系如图4b所示,Gs随着VPD的变化呈二次方关系(R2=0.43,P<0.001),当VPD<1.5kPa时,Gs随VPD的增加而增大,当VPD>1.5kPa时,Gs随VPD的增大有减小趋势,说明高VPD对Gs产生了抑制。有研究认为Gs与VPD呈现负的对数函数关系[36],但影响冠层导度的因素众多,当VPD较小时,VPD的增大并未限制植物气孔活动,而此时Ta和PAR等气象因子对Gs有促进作用,但随着VPD增大到一极值,气孔会因为叶片水势的降低以及蒸腾的增加而闭合,导致Gs减小。

图4 REW>0.4条件下冠层导度与气象因子关系图,误差线为标准差Fig.4 Relationship between canopy conductance(Gs)and climatic variables at REW>0.4,the error bar represent the SD
REW>0.4条件下各因子对生态系统GEP和ET的影响如图5所示,生态系统GEP分别与Ta和PAR呈显著的正相关关系(Ta:R2=0.89,P<0.001;PAR:R2=0.87,P<0.001),随其增加而增大(图5)。生态系统ET和Ta呈显著的正相关关系(R2=0.73,P<0.001),而VPD作为水汽蒸发的驱动力,直接控制着地面的蒸发和植被的蒸腾速率,当VPD<1.5kPa时,ET随着VPD的增加而变大,但当VPD>1.5kPa后,高VPD会抑制冠层导度,导致ET略有降低(图5)。WUE与VPD显示出显著负相关性(图5),这一变化趋势与大多研究结果相似[8,10]。随着VPD的增大,生态系统GEP和ET同时增大,但生态系统ET对VPD的影响更为敏感,因此导致WUE随VPD的增大而减小。但也有研究认为,WUE与VPD的负相关性仅限于温带植被在夏季生长季适用,在较寒冷地区和其他季节,温度和物候对WUE的影响占主导地位[39]。
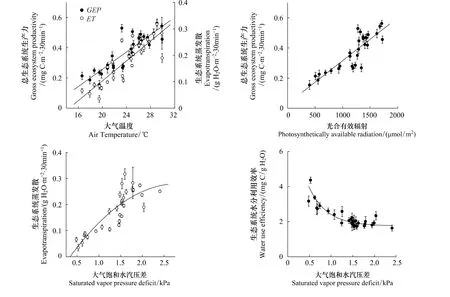
图5 REW>0.4条件下各气象因子对生态系统总生产力,蒸发散和水分利用效率的影响,误差线为标准差Fig.5 The effects of climatic factors on gross ecosystem productivity(GEP),evaportranspiration(ET)and water use efficiency(WUE)at REW>0.4 conditions in the growing season of 2007,the error bar represent the SD
3 结论
通过分析北京大兴区永定河沙地杨树人工林生态系统GEP、ET和WUE的变化情况,讨论了不同土壤水分条件下气象因子和下垫面因素对GEP、ET和WUE的影响。研究发现,即使生态系统受到水分限制,杨树人工林仍然为碳汇。无论是否受到土壤水分胁迫,GEP和ET都显示出较为显著的相关性。
当REW<0.1时,生态系统处于严重水分胁迫状态,土壤水分限制了植物的生理活动,植物受到水分胁迫的影响而关闭气孔,GEP和ET维持在一较低水平,且不随各气象因子的变化发生改变。当生态系统处于土壤水分胁迫期间(0.1<REW<0.4),随着土壤含水量的增加,植物气孔张开,冠层导度逐渐增大,生态系统GEP和ET受到VWC的影响随之迅速增大。未受水分胁迫时(REW>0.4),生态系统GEP和ET主要受到气象因子的影响,并与之呈正相关关系。
较差的土壤保水能力导致土壤水分对ET的影响大于GEP,而气孔对蒸腾的直接控制导致ET对冠层导度的响应快于GEP,因此当生态系统处于土壤水分胁迫期间(0.1<REW<0.4),WUE随着土壤含水量的增加而降低,未受水分胁迫时(REW>0.4),WUE与VPD呈负相关关系。土壤水分胁迫期间,相似的GEP与较高的WUE说明,当VWC处于5.2%—8.8%(0.1<REW<0.4)范围时,土壤水分能够被植物有效利用,达到碳固定对水分消耗的最大效率。
本研究从较小时间尺度对比了不同水分条件下GEP、ET和WUE对各环境因子的响应,并未从不同时间尺度分析各环境因子与碳水通量之间的关系。此外,影响生态系统碳水耦合的因素很多,除了植被内部驱动力气孔行为控制和外部驱动力环境因子影响外,土壤蒸发也对WUE的变化产生很大作用。因此,在未来的研究中,要加强不同时空尺度对比分析,以及土壤蒸发对ET的影响研究,进一步了解杨树人工林生态系统碳水循环机理。
[1]Chinese Forestry Society.National Poplar Commission.Forest resource,timber production and poplar culture in China.In:1st International Conference on the Future of Poplar,Rome,2003.http://www.fao.org/forestry/site/20587/en/.
[2]Sala O E,Parton W J,Joyce L A,Lauenroth W K.Primary production of the central grassland region of the United States.Ecology,1988,69(1):40-45.
[3]Imanuel N M.Desert ecosystems:environment and producers.Annual Review Ecology and Systematicas,1973,4:51-58.
[4]Yu G,Wang Q,Zhuang J.Modeling the water use efficiency of soybean and maize plants under environmental stresses:application of a synthetic model of photosynthesis-transpiration based on stomatal behavior.Journal of Plant Physiology,2004,161(3):308-318.
[5]Baldocchi D D.Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems:past,present and future.Global Change Biology,2003,9(4),479-492.
[6]Barr A G,Black T A,Hogg E H,Griffis T J,Morgenstern K,Kljun N,Theede A,Nesic Z.Climatic controls on the carbon and water balances of a boreal aspen forest,1994—2003.Global Change Biololgy,2006,12(3),1-16.
[7]Veron SV,Paruelo JM,Sala OE,Lauenroth WK.Environmental controls of primary production in agricultural systems of the Argentine Pampas.Ecosystems,2002,5(7):625-635.
[8]Law B E,Waring R H,Anthoni P M.Measurements of gross and net ecosystem productivity and water vapour exchange of a Pinus ponderosa ecosystem and an evaluation of two generalized models.Global Change Biology,2000,6(2):155-168.
[9]Nogueira A,Martinez CA,Ferreira LL,Prado CHBA.Photosynthesis and water use efficiency in twenty tropical tree species of differing succession status in a Brazilian reforestation.Photosynthetica,2004,42(3):351-356.
[10]Tang J,Paul V B,Brent E E,Ankur R D,Kenneth J D,Eileen V C.Sap flux-upscaled canopy transpiration,stomatal conductance,and water use efficiency in an old growth forest in the Great Lakes region of the United States.Geophysical Research,Vol.111,G02009.
[11]Yu G R,Song X,Wang Q F.Water-use efficiency of forest ecosystems in eastern China and its relations to climatic variables.New Phytologist,2008,177(4):927-937.
[12]Kuglitsch F G,Reichstein M,Beer C,Carrara A,Ceulemans R,Granier A,Janssens IA,Koestner B,Lindroth A,Loustau D,Matteucci G,Montagnani L,Moors EJ,Papale D,Pilegaard K,Rambal S,Rebmann C,Schulze ED,Seufert G,Verbeeck H,Vesala T,Aubinet M,Bernhofer C,Foken T,Grunwald T,Heinesch B,Kutsch W,Laurila T,Longdoz B,Miglietta F,Sanz M J,Valentini R.Characterisation of ecosystem water-use efficiency of european forests from eddy covariance measurements,Biogeosciences Discuss,2008,5(6),4481-4519.
[13]Migliavacca M,Meroni M,Manca G,Matteucci G,Montagnani L,Grassi G,Zenone T,Teobaldelli M,Goded I,Colombo R,Seufert G.Seasonal and interannual patterns of carbon and water fluxes of a poplar plantation under peculiar eco-climatic conditions.Agricultural and Forest Meteorology,2009,149(9):1460-1476.
[14]Wei Y,Zhang X D,Jiang Z P,Zhou JX,Tang Y X,Wu L X,Huang L L,Gao SH.Study on the seasonal dynamic of Net Ecosystem Exchange over a poplar plantation of Yueyang city in Hu nan province.Forest Research,2010,23(5):656-665.
[15]Peng Z H,Wang Y,Ren H Q,Sun Q X,Zhou J X.Research on the variation of carbon flux and the relationship of environmental factors and carbon flux of populous forest ecosystem in the reaches of Yangtze River in Anqing.Forest Research,2009,22(2):237-242.
[16]Noormets A(2008).http://www4.ncsu.edu/—anoorme/ECP/[EB/OL].
[17]Webb E K,Pearman G I,Leuning R.Correction of flux measurements for density effects due to heat and water vapour transfer.Quarterly Journal of the Royal Meteorological Society,1980,106(447):85-100.
[18]Lloyd J,Taylor JA.On the temperature dependence of soil respiration.Funcional Ecology,1994,8(3),315-323.
[19]Wen X F,Yu G R,Sun X M,Li QK,Liu Y F,Zhang L M,Ren CY,Fu Y L,Li Z Q.Soil moisture effect on the temperature dependence of ecosystem respiration in a subtropical Pinus plantation of southeastern China.Agricultural and Forest Meteorology,2006,137(3/4):166-175.
[20]Fang X R,Zhang Z Q,Zha T G.Tan J R,Wang X P,Chen J Q,Chen J Q,Sun G.Ecosystem and soil respiration of a poplar plantation on a sandy floodplain in Northern China.Acta Ecologica Sinica,2012,32(8):2401-2409.
[21]Monteith JL.Evaporation and environment.Symposia of the Society for Experimental Biology,1965,19:205-234.
[22]Granier A,Bre'da N,Biron P,Villette S.A lumped water balance model to evaluate duration and intensity of drought constraints in forest stands.Ecological Modelling,1999,116(2):269-283.
[23]Bernier P Y,Bre'da N,Granier A,Raulier F,Mathieu F.Validation of a canopy gas exchange model and derivation of a soil water modifier for transpiration for sugar maple(Acer saccharum Marsh.)using sap flow density measurements.Forest Ecology and Management,2012,163(1):185-196.
[24]Mahrt L.Flux sampling errors for air craft and towers.Journal of Atmospheric and ocean technology,1998,15,416-29.
[25]Wilson K,Goldstein A,Falge E,Aubinet M,Baldocchi D,Berbigier P,Bernhofer C,Ceulemans R,Dolman H,Field C,Grelle A,Ibrom A,Law B E,Kowalski A,Meyers T,Moncrieff J,Monson R,Oechel W,Tenhunen J,Valentini R,Werma S.Energy balance closure at FLUXNET sites.Agricultural and Forest Meteorology,2002,113(1),223-43.
[26]Williams D G,Cable W,Hultine K,Hoedjes J C B,Yepez E A,Simonneaux V,Er-Raki S,Boulet G,De Bruin H A R,Chehbouni A,Hartogensis O K,Timouk F.Evapotranspiration components determined by stable isotope,sap flow and eddy covariance techniques.Agricultural and Forest Meteorology,2004,125(3),241-258.
[27]Law B E,Falge E,Gu L,Baldocchi D D,Bakwin P,Berbigier P,Davis K,Dolman A J,Falk M,Fuentes JD,Goldstein A,Grelle A,Hollinger D,Janssens I,Jarvis P,Jensen N O,Katul G,Mahli Y,Matteucci G,Monson R,Munger W,Oechel W,Olson R,Pilegaard K,Paw U K T,Thorgeirsson H,Valentini R,Verma S,Vesala T,Wilson K,Wlfsy S.Environmental controls over carbon dioxide and water vapor exchange of terrestrial vegetation.Agricultural and Forest METeorology,2002,113(1):97-120.
[28]Huber SC,Rogers H H,Mowry F L.Effects of water stress on photosynthesis and carbon partitioning in soybean[Glycine max(L.)Merr.]plants grown in the field at different CO2levels.Plant Physiology,1984,76(1):244-249.
[29]Nijs I,Impens I,Behaeghe T.Effects of long-term elevated atmospheric CO2concentration on Lolium perenne and Trifolium repens canopies in the course of a terminal drought stress period.Canada Journal of Botony,1989,67(9):2720-2725.
[30]Liang N,Maruyama K.Interactive effects of CO2enrichment and drought stress on gas exchange and water-use efficiency in Alnus firma.Environmental and Experimental Botany,1995,35(3):353-361.
[31]Reichstein M.,Tenhunen JD,Roupsard O,Ourcival JM,Rambal S,Dore S,Valentini R.Ecosystem respiration in two Mediterranean evergreen Holm Oak forests:drought effects and decomposition dynamics.Functional Ecology,2002,16(1):27-39.
[32]Teskey R O,Gholz H L,Cropper J.Influence of climate and fertilization on net photosynthesis of mature slash pine.Tree Physiology,1994,14(11):1215-1227.
[33]Granier A,Reichstein M,Breda N,Janssens IA,Falge E,Ciais P,Grunwald T,Aubinet M,Berbigier P,Bernhofer C,Buchmann N,Facini O,Grassi G,Heinesch B,Ilvesniemi H,Keronen P,Knohl A,Kostner B,Lagergren F,Lindroth A,Longdoz B,Loustau D,Mateus J,Montagnani L,Nys C,Moors E,Papale D,Peiffer M,Pilegaard K,Pita G,Pumpanen J,Rambal S,Rebmann C,Rodrigues A,Seufert G,Tenhunen J,Vesala T,Wang Q.Evidence for soil water control on carbon and water dynamics in European forests during the extremely dry year:2003.Agricultural and Forest Meteorology,2007,143(1/2):123-145.
[34]Song X,Yu G R,Liu Y F,Sun X M,Lin Y M,Wen X F.The variation and effect of environmental factors on Water-use efficiency of subtropical plantation.Science in China Ser.D Earth Sciences,2006,36(supplementary issue I):111-118.
[35]Law B E,Waring R H,Anthoni P M.Measurements of gross and net ecosystem productivity and water vapour exchange of a Pinus ponderosa ecosystem and an evaluation of two generalized models.Global Change Biology,2000,6(2):155-168.
[36]Tang J,Paul V B,Brent E E,Ankur R D,Kenneth J D,Eileen V C.Sap flux-upscaled canopy transpiration,stomatal conductance,and water use efficiency in an old growth forest in the Great Lakes region of the United States.Geophysical Research,Vol.111,G02009.
[37]Dewar R C.A simple model of light and water use evaluated for Pinus radiata,Tree Physiology,1997,17(4):259-265.
[38]Berbigier P,Bonnefond J M,Mellmann P.CO2and water vapour fluxes for two years above Euroflux forest site,Agricultural and Forest Meteorology,2001,108(3):183-197.
[39]Goulden M L,Daube BC,Fan SM,Sutton D J,Bazzaz A,Munger W,Wofsy SC.Physiological responses of a black spruce forest to weather.Journal of Geophysical Research,1997,102:28987-28996.
参考文献:
[14] 魏远,张旭东,江泽平,周金星,汤玉喜,吴立勋,黄玲玲,高升华.湖南岳阳地区杨树人工林生态系统净碳交换季节动态研究.林业科学研究,2010,23(5):656-665.
[15] 彭镇华,王妍,任海青,孙启祥,周金星.安庆杨树林生态系统碳通量及其影响因子研究.林业科学研究,2009,22(2):237-242.
[20] 方显瑞,张志强,查同刚,谭炯锐,王小平,陈俊琦,陈吉泉,孙阁.永定河沿河沙地杨树人工林生态系统呼吸特征.生态学报,2012,32(8),2401-2409.
[34] 宋霞,于贵瑞,刘允芬,孙晓敏,林耀明,温学发.亚热带人工林水分利用效率的季节变化及其环境因子的影响.中国科学:D辑 地球科学,2006,36(增刊I):111-118.
