北京九龙山自然保护区植物群落物种多样性分析
2013-09-14胡淑萍刘鹏举高开通唐小明
胡淑萍,刘鹏举,高开通,唐小明
(中国林业科学研究院 资源信息研究所,北京100091)
物种多样性可以表征一个群落的结构类型、组织水平、发展阶段、稳定程度和生境差异等[1]。一个地区植物群落的构成是这个地区气候、土壤以及人为等多方面因素的综合反映。在同一地区可以有不同的群落类型,即使在同一地区看似相同的群系外貌,其内部结构也可能存在很大差异[2]。由于物种多样性是植被建设和恢复的重要定量指标[3-4],所以阐明群落多样性与相关影响因子之间的关系至关重要。
北京山区由于地域经济开发较早,植被破坏严重,其物种多样性特征一直备受诸多学者的关注,尤其在东灵山、喇叭沟门、百花山、蒲洼自然保护区等已开展过大量关于物种多样性的研究[5-10]。九龙山自然保护区是保护典型暖温带石质山地次生落叶阔叶林生态系统的自然保护区,自然环境复杂、生物多样性丰富是该区的主要特点。本文运用灰色关联法对九龙山自然保护区群落的物种多样性进行分析,旨在研究其群落特征、物种组成及影响物种多样性的主要因子,以期找到提高物种多样性的可控因子,为森林可持续经营提供依据。
1 研究区概况
北京九龙山自然保护区位于北京市西郊,门头沟区东部。地理坐标为东经115°59′—116°06′,北纬39°54′—39°57′;保护区南北宽约6 k m,东西长约10 k m,总面积1 333.4 h m2;平均海拔150~350 m,最高海拔850 m。区内气候属暖温带半湿润季风气候,四季分明,年均温11℃,有效积温3 385~4 210℃,无霜期140 d左右。年降水量600 mm,汛期(6—9月)降水量约占全年的85%。土壤多为淋溶褐土和棕壤,土层厚度60~90 c m,通气透水性较好。植被多由人工林组成,主要乔木树种有黄栌(Cotinus coggygria)、侧柏(Pl atycl adus orientalis)和油松(Pinus tabul aef or mis);灌木树种有荆条 (Vitex negundo var.heterophyll a)、胡 枝 子 (Lespedeza bicolor Turcz)、雀儿舌头(Leptopus chinensis)、孩儿拳头(Grewia biloba var.par vif lor a)等;草本植物主要有东亚唐松草(Thalictr u m minus var.hypoleucu m)、披 针 叶 苔 草 (Carex l anceol ata)、求 米 草(Oplismenus undul atif olius)、蝎子草(Sedu m spectabile Boreau)、绒毛绣线菊(Spir aea dasyantha)等。
2 研究方法
2.1 样地设置与调查
根据地形因子与群落特征设置8块20 m×20 m的样地,对每块样地内的乔木进行每木检尺,记录种名、胸径、树高、冠幅、健康状况等因子,同时记录样地的海拔、坡度、坡向等生境因子。在每块样地内设置5个2 m×2 m的灌木样方和10个1 m×1 m的草本样方,分别记录灌木和草本的种类、高度和盖度等因子(表1)。

表1 调查样地基本特征
2.2 数据分析与计算
2.2.1 物种多样性计算方法
(1)物种重要值计算

(2)物种多样性测度
物种多样性计算选用Si mpson优势度指数、Shannon-Wiener多样性指数、Margalef丰富度指数和Piel ou均匀度指数,其计算公式为:
Si mpson优势度指数(C):

式中:Pi——种i的相对重要值,Pi=ni/N,ni——群落中某一层次第i个物种的重要值;N——该层次所有物种重要值之和;S——种i所在层的总物种数。
Shannon-Wiener多样性指数(H):

式中:Pi,S 含义同式(3)。
Mar galef丰富度指数(R):
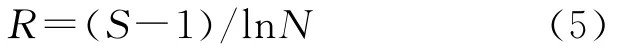
式中:S——群落中的总物种数;N——群落中全部种的总个体数。
Pielou均匀度指数(J):

式中:H——Shannon-Wiener多样性指数;S——群落中的总物种数。
群落总体多样性:

式中:Wi——群落第i个生长型多样性指数的加权参数;Di——第i个生长型的多样性指数(i=1:乔木层;2:灌木层;3:草本层)。加权参数的计算公式为[11-12]:
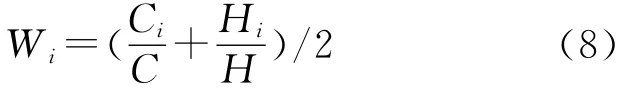
式中:C——群落的总盖度(C=∑Ci);H——群落各生长型的平均高度(H=∑Hi);Ci——第i个生长型的盖度;Hi——第i个生长型平均高度。经计算,乔灌草三层的权重分别为0.85,0.12和0.03。
2.2.2 灰色关联分析法 灰色关联分析是灰色理论中的一种分析方法,用来分析系统中母因素与子因素的关系密切程度,从而判断引起该系统发展的主要和次要因素[13-14]。灰色关联分析是依据各因素数列曲线形状的接近程度做发展态势的分析,简言之是指在系统发展过程中,如果两个因素变化的态势是一致的,即同步变化程度较高,则可以认为两者关联较大;反之,则两者关联度较小。
(1)确定参考序列和比较序列。
参考序列又称母序列,记为Yi,i∈M={1,2,…,m},一般选取主研究对象为参考序列。比较序列又称子序列,记为Xj,j∈N={1,2,…,n},通常选取影响主要研究对象的因素序列为比较序列。序列表示为:

其中,k∈L={1,2,…,l}。
(2)计算关联系数。

其中ρ为分辨系数,一般取0到1,本研究取值为0.5。
(3)计算关联度。
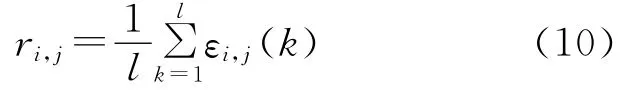
其中i∈M,j∈N,k∈L。
3 结果与分析
3.1 群落结构特征
研究区建群种主要为黄栌和侧柏(表2),其中黄栌林分布于东北和西北坡向,物种数在21~41之间,林分密度在1 325~1 775株/h m2之间;侧柏林分布在东南、西北和西南坡向,物种数为19~39,林分密度为875~1 675株/h m2。
各样地主要物种重要值如表2所示。样地1共有乔木4种,其中黄栌为建群种(0.936 2),灌木层以荆条(0.469 0)、胡枝子(0.175 3)、杭子梢(0.107 5)为主,草本层以披针叶苔草(0.183 5)、求米草(0.119 3)和东亚唐松草(0.113 1)为主。样地2以黄栌(0.460 7)和油松(0.425 4)为建群种,灌木层共有植物12种,其中荆条(0.467 5)、多花胡枝子(0.111 7)、胡枝子(0.103 1)占到总重要值的68%,草本层共有植物24种,其中求米草(0.113 1)、披针叶苔草(0.098 9)、北 京 隐 子 草 (0.084 5)占 总 重 要 值 的30%。样地3的建群种是黄栌(0.423 3)和油松(0.395 1),灌木层重要值排序前3位的依次是荆条(0.500 2)、胡枝子(0.259 9)和小叶鼠李(0.080 7),草本层重要值排序前3位的是求米草(0.245 6)、披针叶苔草(0.181 2)和北京堇菜(0.164 0)。样地4乔木层由黄栌(0.892 7)和油松(0.107 3)组成,灌木层中荆条(0.425 3)、孩儿拳头(0.123 0)、雀儿舌头(0.106 4)占据优势,草本层中披针叶苔草(0.176 1)、求米草(0.136 0)、秋苦荬(0.120 4)占据优势。样地5乔木层由侧柏(0.664 0)和元宝枫(0.336 0)组成,灌木层荆条(0.882 5)、胡枝子(0.086 2)、红花锦鸡儿(0.018 7)占到总重要值的99%,草本层线叶猪秧秧(0.334 6)、北京隐子草(0.196 8)、蝎子草(0.131 0)占到总重要值的66%。样地6建群种为侧柏(0.694 8),灌木层以荆条(0.661 4)、多花胡枝子(0.142 3)、孩儿拳头为主(0.060 7),草本层以北京隐子草(0.170 3)、铁杆蒿(0.161 6)、狗尾草(0.116 6)为主。样地7以侧柏(0.637 0)和黄栌(0.215 1)为建群种,灌木层共有11种植物,其中荆条(0.437 0)、多花胡枝子(0.151 1)、酸枣(0.105 4)占总重要值的69%,草本层共有18种植物,其中披针叶苔草(0.166 9)、秋苦荬(0.158 1)、北京隐子草(0.102 6)占总重要值的43%。样 地 8 乔 木 层 由 黄 栌 (0.515 3)、侧 柏(0.468 3)、油松(0.016 3)3种植物组成,灌木层重要值排序前3位的依次是荆条(0.431 4)、胡枝子(0.260 0)、孩儿拳头(0.098 5),草本层重要值排序前3位的是披针叶苔草(0.175 8)、求米草(0.100 5)、绒毛绣线菊(0.075 2)。
3.2 群落物种多样性比较
表3显示的是各样地不同生长型的多样性指数。乔木层中,样地3和样地2的Si mpson指数均高于0.6,而样地4和样地1的Si mpson指数低于0.2;样地3的Shannon-Wiener指数最大,为1.260 5,这与该样地乔木种类多、且分布均匀有关;样地7和样地3的Mar galef指数较大,这与两个样地乔木层树种较多的现状相符;样地5中侧柏和元宝枫的分布较均匀,所以Piel ou指数最大。
灌木层中,样地4的Si mpson指数和Shannon-Wiener指数最大,分别为0.773 1和1.973 4;样地5的Si mpson指数和Shannon-Wiener指数最小,仅为0.213 2和0.451 1;样地4灌木层物种数为18种,远高于其他样地,所以Mar galef指数最大,为3.857 7;灌木层各样地Pielou指数多集中在0.56~0.79之间,仅样地5的Piel ou指数为0.325 4。

表2 各样地主要物种的重要值

表3 各样地多样性指数
草本层中,各样地的Si mpson指数在0.816 4~0.936 2之间;Shannon-Wiener指数在1.978 3~2.936 2之间;样地8和样地2的Margalef指数较大,这与其草本层物种数均高于其他样地有关;各样地草本层Pielou指数分布在0.792 0~0.918 0之间。
群落总体多样性变化规律显示,各样地Si mpson指数大小依次为:样地3>样地2>样地7>样地8>样地6>样地5>样地4>样地1;Shannon-Wiener指数大小依次为:样地3>样地2>样地7>样地6>样地8>样地5>样地4>样地1;Mar galef指数大小依次为:样地7>样地2>样地3>样地6>样地1>样地8>样地4>样地5;Pielou指数大小依次为:样地5>样地3>样地8>样地2>样地6>样地7>样地4>样地1。
3.3 群落多样性的物种多样性分析
依据灰色系统理论的要求,在分析各因子对生物多样性的影响时,将Si mpson指数、Shannon-Wiener指数、Margalef指数、Pielou指数作为参考序列 ,密度、物种数、枯落物厚度、海拔等6个因子作为比较序列Yj。由于原始数据中各个因子的物理意义不同,为保证各因子具有等效性和同序性,对原始数据采用标准差法进行无量纲化处理,处理结果如表4所示。

表4 无量纲化多样性指数及相关影响因子
表5显示的是群落多样性指数与相关影响因子的关联系数及排序。表5表明,各因子与群落多样性指数的灰色关联度均较高,且不同的多样性指数与影响因子之间的相关性与排序各不相同。Si mpson指数受坡向影响最大,其次为坡度、海拔等。Shannon-Wiener指数与坡向的关联系数最大,为0.845 3,而与其他影响因子的关联度在0.623 1~0.676 0之间。Mar galef指数与坡向的关联系数达0.777 3,其次分别为枯落物厚度(0.688 9)、物种数(0.684 2)、坡度(0.649 2)、密度(0.624 6)和海拔(0.606 8)。Piel ou指数与影响因子的相关性排序依次为:坡度>枯落物厚度>海拔>坡向>密度>物种数。
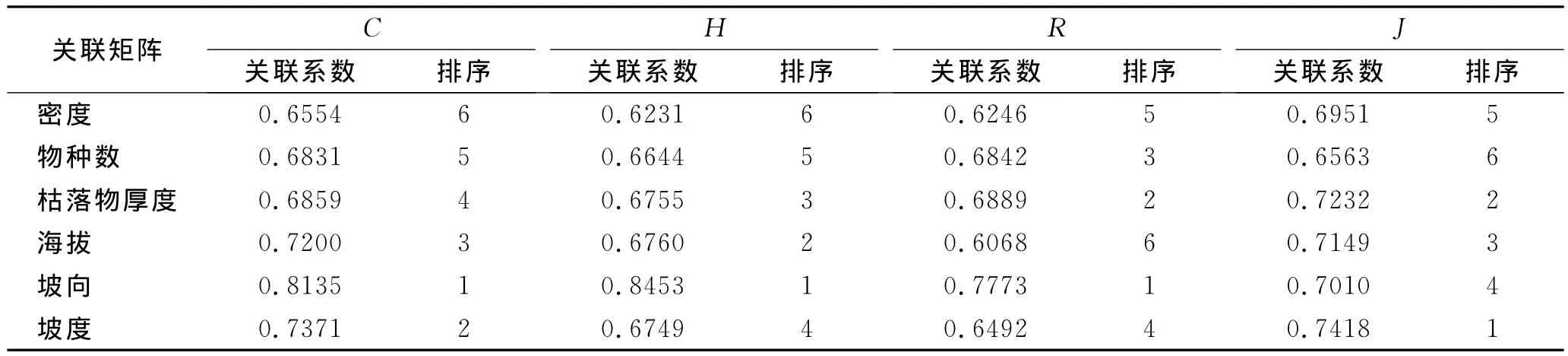
表5 多样性指数与相关影响因子的关联度
4 结论
(1)研究区建群种为黄栌和侧柏,其中黄栌林分布于东北和西北坡向,侧柏林分布在东南、西北和西南坡向。各样地林分密度变化较大,但林下灌木层均以荆条为主,其重要值介于0.425 3~0.882 5之间,其次为胡枝子、小叶鼠李、孩儿拳头等,而草本层中披针叶苔草、求米草、线叶猪秧秧、北京隐子草的重要值较大。
(2)各样地物种多样性变化多呈草本层多样性最高,灌木层次之,乔木层最低的趋势。群落总体多样性显示,样地3的Si mpson指数和Shannon-Wiener指数最大,样地7的 Mar galef指数最高,样地5的Pielou指数最大。
(3)群落多样性指数与各影响因子之间的灰色关联系数均较高,其中Si mpson指数、Shannon-Wiener指数和Mar galef指数受坡向影响显著,Pielou指数受坡度影响最大,其次为枯落物厚度、海拔和坡向。
(4)目前多采用相关分析、回归分析等方法研究环境因子与物种多样性之间的关系,但由于此类方法需要大量的样本数据,且要求样本具有一定的统计规律,服从典型的概率分布。本研究由于样本数量少,选用灰色关联分析作为研究方法,充分发挥其所需样本量少、方法简便、信息量大等优点,对研究环境因子与物种多样性关系具有一定的参考意义。
(5)由于本研究中的8块样地距离较为接近,所以视各样地土壤条件相对均一。人工林物种多样性变化受多种因素影响,本研究主要分析了环境因子与林分基本状况对群落物种多样性的影响,而并未考虑干扰等影响因子,不足之处需要在下一步工作中继续完善。
[1] 阎海平,谭笑,孙向阳,等.北京西山人工林群落物种多样性的研究[J].北京林业大学学报,2001,23(2):16-19.
[2] 卢宝明,邢韶华,崔国发,等.北京山地植物群落的物种多样性比较[J].北京林业大学学报,2010,32(Z1):36-44.
[3] Ruiz-Jaen M C,Aode T M.Restoration success:how is it being measured [J].Restoration Ecology,2005,13(3):569-577.
[4] Zerbe S,Kreyer D.Introduction to special section on‘ecosystem restoration and biodiversity:how to assess and measure biological diversity’[J].Restoration Ecology,2006,14(1):103-104.
[5] 马克平,刘灿然,于顺利,等.北京东灵山地区植物群落多样性的研究[J].生态学报,1997,17(6):574-583.
[6] 张育新,马克明,祁建,等.北京东灵山辽东栎林植物物种多样性的多尺度分析[J].生态学报,2009,29(5):2179-2185.
[7] 崔国发,成克武,申国珍,等.北京喇叭沟门林区森林植被现状及分类[J].北京林业大学学报,2000,22(4):46-51.
[8] 成克武.北京喇叭沟门林区植物多样性及其保护研究[D].北京:北京林业大学,2002.
[9] 郑景明,赵秀海,张春雨.北京百花山森林群落的结构多样性研究[J].北京林业大学学报,2007,29(1):7-11.
[10] 陈维川,赵勃,邢韶华,等.北京蒲洼自然保护区植物群落的优先保护顺序[J].东北林业大学学报,2005,33(5):62-64.
[11] 高贤明,黄建辉,万师强,等.秦岭太白山弃耕地植物群落演替的生态学研究:Ⅱ.演替系列的α多样性特征[J].生态学报,1997,17(6):619-625.
[12] 高宝嘉,张执中,李镇宇.封山育林对植物群落结构及多样性的影响[J].北京林业大学学报,1992,14(2):46-53.
[13] 邓聚龙.灰色系统理论教程[M].武汉:华中理工大学出版社,1990.
[14] 蔡文春,杨德刚.新疆耕地和粮食灰色关联分析[J].干旱区资源与环境,2007,21(11):54-58.
