黄土丘陵沟壑区不同侵蚀环境下幼苗库动态变化特征
2013-09-14焦菊英王志杰杜华栋
苏 嫄,焦菊英,王志杰,杜华栋
(1.西北农林科技大学 资源环境学院,陕西 杨凌712100;2.中国科学院 水利部水土保持研究所,陕西 杨凌712100;3.西北农林科技大学 水土保持研究所,陕西 杨凌712100)
陕北黄土丘陵沟壑区由于降水稀少、气候干旱,加之长期的过度农耕和肆意放牧,导致自然植被遭到严重破坏,土壤侵蚀极为严重[1],成为我国生态环境最为脆弱的地区之一,生态系统亟待修复[2-3]。植被作为重要的生态因子,是防止生态退化的物质基础[4],也是控制水土流失、改善生态环境的有效措施和保证,在自然生态环境中占有极其重要的地位[5]。因此,植被恢复是黄土高原遏制土地退化,促进退化生态系统恢复的关键因素和有效途径[6-8]。
幼苗库(seedling bank)是指一定面积的样地中小于某一高度的所有植物幼苗的总和[9],是植被群落的一部分,也是植被恢复演替的基础。幼苗库中的幼苗通常会通过参与植被群落的自然更新来影响群落中成年植物的分布和丰富度,以及地上植被群落的组成、结构、动态变化和物种多样性维持[10-11]。可见,幼苗库是植被动态的重要制约因素,影响着生态系统的抗干扰能力和恢复能力,对植物物种和群落的恢复与保护具有重要作用。因此,研究幼苗库特征及其影响因素,在一定程度上可为人为干扰植被的恢复提供理论依据,对黄土高原的植被重建具有一定的指导意义。
目前对幼苗库已有大量研究并取得了重要进展,主要集中在幼苗更新特性及影响因子[12-14]、幼苗种群特征与分布格局[15]、幼苗库与种子库的关系[9]、幼苗建植[16-17]以及幼苗与自然植被的关系[18]等方面。但这些研究往往是一种定性的描述或是静态的定量比较,关于幼苗库特征的动态变化研究还比较少,而这又是研究幼苗更新能力急需解决的问题。为此,本研究通过对黄土丘陵沟壑区典型小流域自然恢复坡面5种侵蚀环境下的15个样地进行定点跟踪观测,分析不同侵蚀环境下幼苗库的动态变化特征,探讨影响幼苗存活的关键影响因子,以期为黄土丘陵沟壑区退化生态系统恢复和生态环境保护提供科学依据。
1 材料与方法
1.1 研究区概况
研究区位于黄土丘陵沟壑区的典型区安塞县纸坊沟和宋家沟流域。安塞县(105°51′44″—109°26′18″E,36°22′40″—36°32′16″N),海拔997~1 731 m,南北长92 k m,东西宽36 k m,土地总面积2 950.2 k m2,境内地形复杂,梁峁连绵,沟壑纵横。属暖温带半干旱气候区,年平均降水量500 mm左右,降雨多集中在7—9月份,占全年降雨总量的63%,多以暴雨形式出现,易造成水土流失[19],土壤类型主要是黄绵土,土质疏松,抗蚀抗冲性差,水土流失严重,耕作层养分含量少,土地贫瘠[20]。在植被分区上属暖温带森林草原带,区内植被破坏严重,天然森林较少,覆盖率为17.7%[21],人工林以刺槐(Robinia psendoacacia)、小叶杨(Popul us si monii)等为主,灌丛主要有柠条(Car agana Korshinskii)和沙棘(Hip pophae r hamnoides)等人工灌丛以及黄刺玫(Rosa xanthina)、虎榛子(Ostr yopsis davidiana)和狼牙刺(Sophor a viciif olia)等天然灌丛[22],草本植物群落主要为猪毛蒿(Artemisia scoparia)、铁杆蒿(Artemisia g melinii)、茭蒿(Artemisia gir al dii)、长芒草(Stipa bungeana)、白羊草(Bothriochloa ischaemu m)、大针茅(Stipa gr andis)、达乌里胡枝子(Lespedeza daurica)等[23]。
1.2 研究方法
1.2.1 样地选择与采样 于2011年10月在安塞县纸坊沟和宋家沟流域选择3个具有代表性的自然恢复梁峁(纸坊沟1个,宋家沟2个),按阳沟坡、阳峁坡、峁顶、阴峁坡、阴沟坡5种不同的侵蚀环境选择调查样地(50 m×50 m)(图1),每种环境设3个重复,共选择固定样地15个,每个样地设置3个2 m×2 m的重复大样方(共45个),并在大样方内按对角线设置3个50 c m×50 c m的小样方(共135个),对小样方的幼苗库特征进行逐月跟踪调查同时进行土壤水分测定。于2012年8月份调查样地的基本情况(表1),包括海拔、坡度、地上植被的盖度、群落类型等,调查样地的养分及水分含量详见表2。
幼苗库跟踪调查于2012年4—11月份的月初进行,每月一次。幼苗库调查中详细记录幼苗的物种组成、数量、高度、生长状况以及死亡情况等。为区分不同调查时间幼苗存活与萌发的个数,调查时用不同颜色的牙签对各月幼苗进行标记。
土壤水分测定采用土钻法,每个样方设置3个重复,取样深度为60 c m,第1层为0—5 c m,以后每隔10 c m一层,共7层,分别取样测定,采用烘干法求其土壤含水量,计算公式为:土壤含水量=(土壤鲜质量-土壤干质量)/土壤干质量×100%。
2012年安塞降雨资料由中国科学院安塞水土保持综合试验站提供。
1.2.2 数据处理 利用Excel 2007软件处理数据,并用SPSS 17.0软件对不同侵蚀环境间的幼苗物种数、幼苗密度、物种多样性指数和丰富度指数进行单因素方差分析(One-Way ANOVA)和LSD检验,对土壤水分、土壤养分、降雨量与幼苗库特征的关系进行相关分析(Bivariate Correlate),显著水平为p<0.05。
Shannon-wiener多样性指数[24]:

Mar galef丰富度指数:Dma=(S-1)/l n N
式中:Pi——第i种个体数占总个体数的比例;S——物种总数;N——所有物种个体总数。

图1 样地分布状况图2 不同侵蚀环境下幼苗物种数动态变化

表1 样地基本情况

表2 不同侵蚀环境下土壤养分与水分含量
2 结果与分析
2.1 不同侵蚀环境下幼苗库的物种组成
15个样地4—11月份共鉴定出幼苗52个物种,隶属23科,其中菊科(Compositae)、豆科(Legu-minoseae)和禾本科(Gr a mineae)这3科的物种占所有物种的50%以上。生长型表现为多年生草本最多(29种),占到总物种的56%,一两年生草本次之(16种),占31%,灌木(5种),占10%,乔木、藤本最少,各1种,分别仅占2%。
不同侵蚀环境下的幼苗物种组成存在差异(表3)。在阳沟坡,年内共记录幼苗29种,其中阿尔泰狗娃花 (11.71%)、菊 叶 委 陵菜 (30.05%)、猪毛 蒿(16.84%)3个物种的幼苗占到该侵蚀环境下幼苗总数的一半以上;阳梁峁坡上共记录34个物种,菊叶委陵菜(24.55%)、长芒草(13.38%)、阴行草(11.73%)、猪毛蒿(10.30%)4个物种占幼苗总数的60%左右;峁顶上共记录了35个物种,其中臭蒿(23.79%)、猪毛蒿(21.71%)、达乌里胡枝子(15.93%)这3个物种所占比例较大;在阴梁峁坡共有46个物种,獐牙菜(19.20%)、铁杆蒿(13.92%)、山苦荬(9.78%)、阴行草(9.40%)4个物种占幼苗总数的一半以上;阴沟坡的幼苗物种较多,共记录47种,其中异叶败酱(15.34%)、长芒草(14.28%)、山苦荬(11.83%)、野菊花(8.18%)、铁杆蒿(5.99%)这4个物种所占幼苗总数的比例较高。对不同侵蚀环境下幼苗物种数进行单因素方差分析得出:阴峁坡和阴沟坡与阳峁坡差异显著(p<0.05),与阳沟坡差异极显著(p=0.001),峁顶与阳沟坡差异显著(p=0.043),与其他4种侵蚀环境均无显著差异(p>0.05)。对幼苗物种数与土壤水分、土壤养分含量进行相关分析可知,幼苗物种数与土壤水分含量呈显著正相关(R=0.887,p=0.048),与土壤养分虽未达到显著相关水平(p>0.05),但幼苗物种数随土壤有机质、全N、速效K含量的增加而增大(R 分别为0.482,0.377,0.513)。另外,地上植被物种组成不同导致不同侵蚀环境下幼苗库物种组成存在差异。可见,侵蚀环境不同,由于土壤水分、养分条件(表2)、地上植被物种组成(表1)不同,幼苗物种数、物种组成存在很大差异。
不同月份之间,幼苗物种数变化趋势表现为4月初较小,平均物种数为10,其中菊科5种:山苦荬、猪毛蒿、铁杆蒿、臭蒿、阿尔泰狗娃花,蔷薇科1种:菊叶委陵菜,唇形科1种:香青兰,禾本科1种:长芒草,亚麻科1种:亚麻,龙胆科1种:獐牙菜;5月初迅速增大,增加的物种主要有小蓟(Cirsium setosum)、牦牛儿苗、火绒草(Leontopodiu m al pinu m)、白羊草、茭蒿等;6月初再次下降,4月、5月份出苗的白羊草、茭蒿、狭叶米口袋、鸦葱、二裂委陵菜这5个物种的幼苗全部死亡;7—9月逐渐增大,物种数达到高峰,主要有达乌里胡枝子、斑种草(Bothriosper mu m chinense)、地黄(Rehmannia gl utinosa)、地锦、狗尾草、尖叶胡枝子、猪毛菜(Salsol a collina)等物种不断出现;10月初和11月中旬又有一定的降低,但高于4月初和6月初(图2)。可见,不同侵蚀环境下幼苗物种组成年内动态变化明显(p=0.001),且物种数呈现出双峰型动态特征,最大值出现在第二个峰值处,即9月初。这与2012年安塞降雨量及其在年内的分配特点(图3)密切相关。
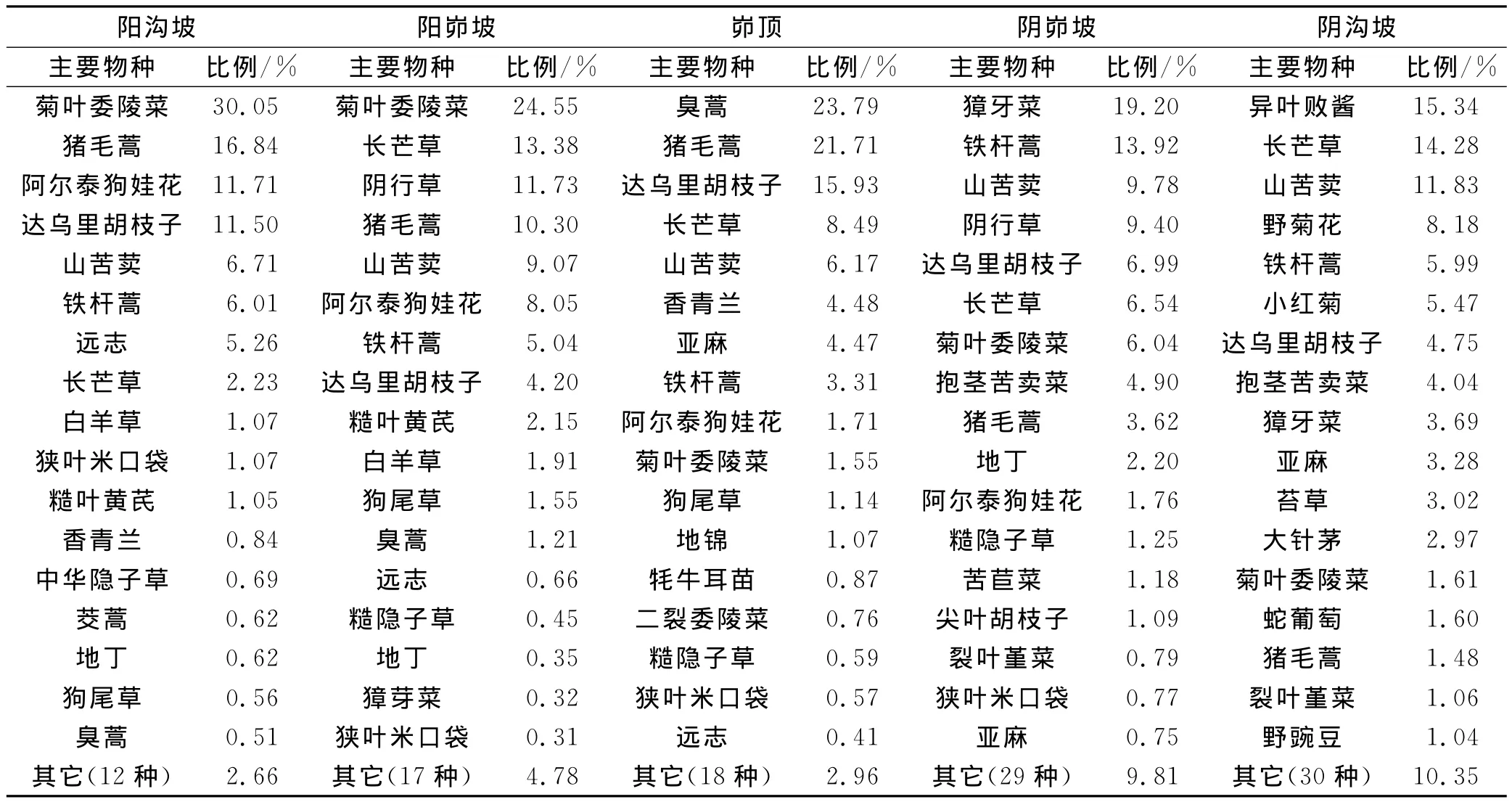
表3 不同侵蚀环境下幼苗主要物种及其所占幼苗总数的比例
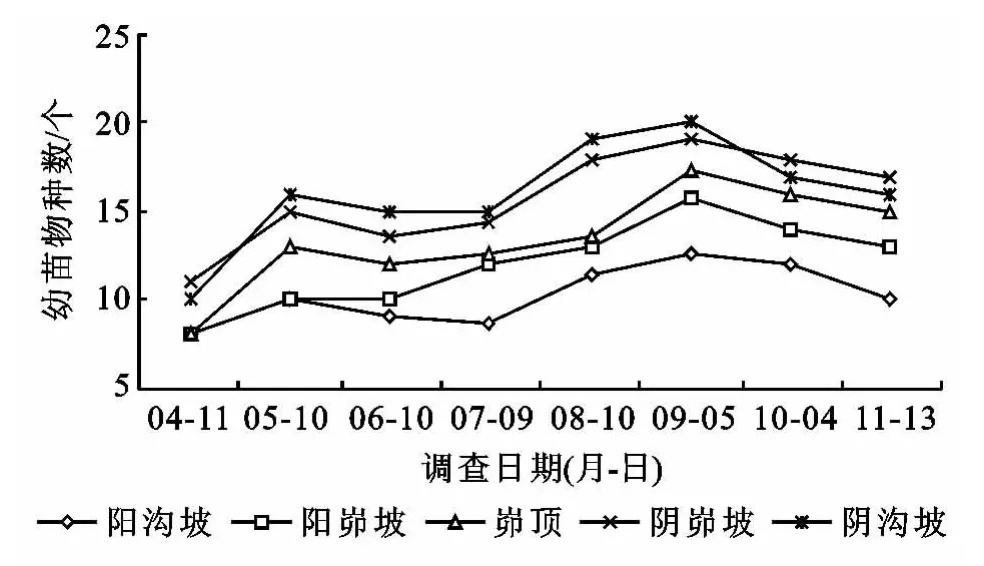
图1 样地分布状况图2 不同侵蚀环境下幼苗物种数动态变化

图3 2012年安塞降雨量动态变化
2.2 不同侵蚀环境下幼苗密度的动态变化
5种不同侵蚀环境下幼苗密度动态变化(图4)表明,不同侵蚀环境之间的幼苗密度存在差异。总体上表现为阴峁坡最大,年内平均值高达78个/m2,与阳沟坡(31个/m2)、阳峁坡(46个/m2)和阴沟坡(48个/m2)差异极显著(p<0.01),而与峁顶(74个/m2)差异不显著(p=0.796);阳沟坡最小且与其他4种侵蚀环境差异显著(p<0.05)。这是土壤水分、土壤养分、侵蚀类型、侵蚀程度综合作用的结果。如阴峁坡不仅土壤水分养分状况较好,而且侵蚀主要是以面蚀和细沟侵蚀为主,侵蚀相对较弱,有利于幼苗生长,故其密度较高。
从不同月份来看,不同侵蚀环境下幼苗密度年内动态变化总体上表现为先升后降的趋势,4月初的幼苗密度较小,之后逐月增大,7月初幼苗密度达到最大值(但阴峁坡6月初幼苗密度最大),之后又逐月减小,至11月中旬幼苗密度又处于低值且低于4月初。最大值(7月初)与最小值(11月中旬)相差较大,其中,阳沟坡相差31个/m2,阳峁坡相差46个/m2,峁顶相差73个/m2,阴峁坡相差78个/m2,阴沟坡相差58个/m2。说明不同侵蚀环境下幼苗密度具有明显的月动态变化特征(p=0.005)。由降雨量与幼苗密度的相关性(表4)可知,幼苗密度与降雨量正相关系数较高,且在阳沟坡和阴沟坡显著相关(p<0.05)。可见,幼苗密度年内动态变化特征与研究区降雨量年内分配特点关系密切。

图4 不同侵蚀环境下幼苗密度动态变化
2.3 不同侵蚀环境下幼苗物种多样性的动态变化
不同侵蚀环境下幼苗物种多样性指数和丰富度指数动态变化如图5所示。不同侵蚀环境之间,幼苗物种多样性指数和丰富度指数均表现为阴峁坡或阴沟坡>峁顶或阳峁坡>阳沟坡,其中阴峁坡和阴沟坡与阳沟坡差异极显著(p<0.01),与其他侵蚀环境之间差异均不显著(p>0.05)。对幼苗物种多样性与土壤水分、养分进行相关分析可知,随土壤水分、土壤有机质、全N、速效K含量的增加,幼苗物种多样性和丰富度指数变大,且与土壤水分呈显著正相关(R=0.878,p=0.049)。可见,土壤水分、养分对幼苗物种多样性有一定的影响。物种多样性与侵蚀环境中侵蚀类型、侵蚀程度以及地上植被也有关系。如阳沟坡坡面陡峭,植被盖度低,侵蚀较为严重,以细沟和浅沟侵蚀为主,该环境不利于幼苗的生长,幼苗不仅萌发少,死亡也较为严重,故幼苗物种较为单一。阴坡地上植被物种种类较多,能为幼苗萌发提供丰富的种源,且土壤水分含量高,促进了种子萌发、幼苗生长,故幼苗也较为丰富多样。
从不同月份来看,不同侵蚀环境下幼苗物种多样性指数和丰富度指数都表现为4月初较小,分别变化在0.42~0.64和1.21~1.60之间,5月初有一定的增大,6月初再次降低,随后呈波动性增长,9月初达到最大值,分别变化在0.68~0.94和2.11~2.97之间,之后又逐渐下降,这与幼苗物种数的动态变化趋势基本一致。由此可见,不同侵蚀环境下幼苗物种多样性和丰富度年内变化呈现明显的规律性变化趋势(p<0.001),即先波动性增大随后降低。且对幼苗物种多样性和丰富度指数动态变化与降雨量的相关性进行分析(表4)可知,幼苗物种多样性和丰富度指数随降雨量的增加而增加,且多样性指数在阴沟坡呈显著正相关(p=0.024)。说明雨季时由于降水多,利于种子的萌发和幼苗的生长,幼苗物种数增多,多样性水平较高;而雨季过后,温度降低,降雨量减少,不仅不利于种子萌发,还会导致大量幼苗死亡,致使幼苗物种数减少,多样性水平降低。
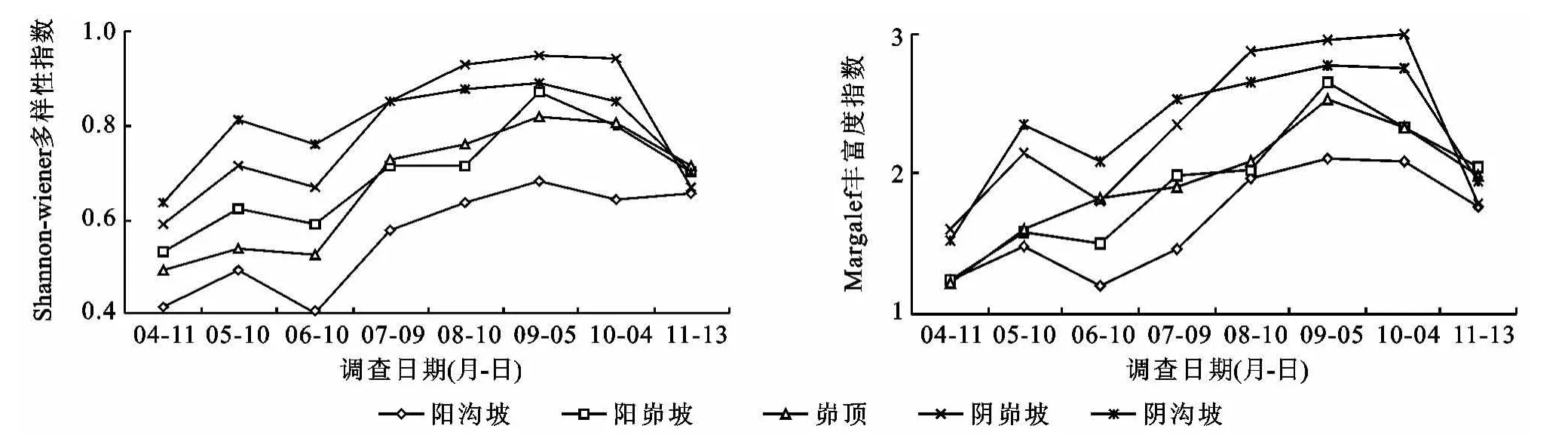
图5 不同侵蚀环境下幼苗物种多样性和丰富度动态变化
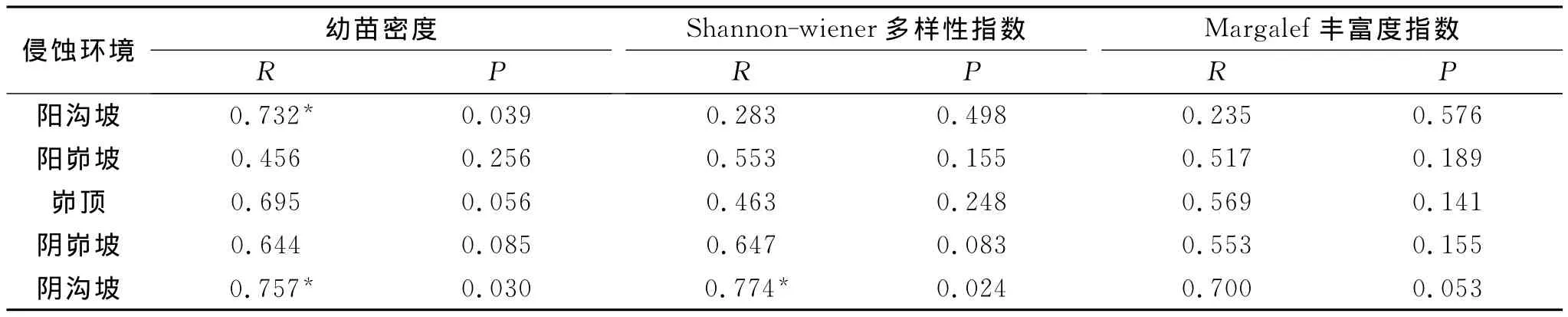
表4 降雨量与幼苗库特征动态变化的相关系数(n=5)
2.4 不同侵蚀环境下幼苗建植特征
由幼苗年内动态变化情况可知,不同侵蚀环境下能够成功定居的幼苗主要物种存在差异。阳沟坡的铁杆蒿、菊叶委陵菜、猪毛蒿、阿尔泰狗娃花,阳峁坡的铁杆蒿、阿尔泰狗娃花、长芒草、菊叶委陵菜,峁顶有铁杆蒿、长芒草、山苦荬、亚麻、阿尔泰狗娃花,阴峁坡的菊叶委陵菜和阴行草,阴沟坡的铁杆蒿、长芒草、阿尔泰狗娃花、抱茎苦荬菜,这些物种的部分幼苗在年内经过雨季和初雪后能够成功定居。可见,铁杆蒿、长芒草、菊叶委陵菜、阿尔泰狗娃花等物种在土壤侵蚀严重的黄土丘陵沟壑区易于存活建植,对地上植被更新和恢复有一定的作用。
3 结论与讨论
本研究对峁坡5种不同侵蚀环境下的幼苗物种组成、幼苗密度、物种多样性指数、丰富度指数动态变化进行了分析,主要得到以下结论:
(1)15个样地4—11月份共调查幼苗52个物种,属23个科,主要由菊科、豆科和禾本科这3科物种组成。生活型方面,多年生草本最多,其次是一两年生草本物种,乔灌木及藤本最少。这与许多学者对该地区地上植被的研究结果基本一致[2,25-26]。不同侵蚀环境间,从阴沟坡到阴峁坡、峁顶、阳峁坡、阳沟坡,幼苗物种数依次减少,且幼苗主要物种及所占幼苗总数的比例也存在差异。这是土壤水分、土壤养分、地上植被群落组成共同作用的结果。阴沟坡不仅土壤水分条件好[27-28],土壤养分含量也相对较高,促进了种子的萌发和幼苗生长,故幼苗物种数较大。阳沟坡虽然土壤养分含量较高,但光照强烈,地面蒸发大[29],土壤水分含量较低[27],且植被盖度低,侵蚀较为严重,导致幼苗大量死亡。不同月份间,幼苗物种数表现为4月初较小,随后波动性增大,9月初达到峰值,10月初和11月中旬又有一定程度的降低,但高于4月初和6月初。这种变化主要是受2012年安塞降雨量及其在年内的分配的影响,同时与地下种子库中种子量大小和种类多少的年际间变化也有一定的关系。
(2)不同侵蚀环境间幼苗密度差异明显,阴峁坡由于土壤养分、水分条件较好,侵蚀相对较弱,幼苗生长状况较好,幼苗密度最大,阳沟坡不仅坡面陡峭,土壤水分含量较低[27],且以细沟和浅沟侵蚀为主,侵蚀严重,不利于幼苗生长,故幼苗密度低。幼苗密度在年内随时间变化具有明显的先升后降的特点。这与降水量及其年内分配特点关系密切,根据2012年安塞的降雨资料,4,5月份降水少,不利于种子的萌发,6,7月份降水逐渐增多幼苗密度达到最大,7月份之后降水减少,幼苗生长受到一定的限制,故密度出现下降趋势。另外,幼苗密度的动态变化可能与温度和地上植被也有一定的关系,4,5月份温度低,不利于种子的萌发,6—8月份温度逐渐升高,但有研究表明,适宜的温度有利于种子的萌发和幼苗生长[30],8月份不仅降水少,同时地上植被生长旺盛,温度高,蒸腾作用强[31],竞争作用剧烈[30],这些都不利于种子的萌发和幼苗生长,导致幼苗密度反而降低。因为本研究中未就温度和地上植被对幼苗动态变化的影响作用进行具体调查,这一方面还有待今后进一步的研究和探讨。
(3)不同侵蚀环境间幼苗物种多样性和丰富度均表现为阴沟坡>阴峁坡>峁顶(阳峁坡)>阳沟坡,与土壤水分含量的变化趋势基本一致。可见,不同侵蚀环境间土壤水分与幼苗物种多样性关系密切。王凯博等[32]的研究也表明,土壤水分条件较差,植被的物种多样性降低。其次,土壤养分、侵蚀类型、侵蚀程度以及地上植被对其都有很大的影响。阴沟坡土壤有机质、全N、速效K含量高[29],能促进幼苗生长,且地上植被物种种类多,可以为幼苗萌发提供丰富的种源[33]。阴峁坡虽然养分含量稍低于阳沟坡,但以面蚀和细沟侵蚀为主,侵蚀相对微弱,对幼苗的破坏作用较小,故阴沟坡和阴峁坡的幼苗物种较为丰富多样。幼苗物种多样性指数和丰富度指数随时间的变化趋势与幼苗物种数相似,即先波动性增大,然后减小。这不仅与降雨量的分配特点密切相关,同时也可能受物种物候期的影响。4—7月份随着降水的增多,幼苗物种数逐渐增加,8月、9月份降水减少,光照强烈,蒸腾作用大,土壤水分含量较低,幼苗难以存活,但也有新的物种不断出现,如斑种草、地黄、地锦、狗尾草、獐牙菜、尖叶胡枝子、猪毛菜等,这些物种主要是在雨季出苗,因而幼苗密度较小,物种数、物种多样性、丰富度指数反而增大。至10月初之后,降水进一步减少,温度降低,幼苗不仅萌发少,甚至有一些物种的幼苗发生死亡,幼苗物种逐渐变得单一。
(4)本研究中对不同侵蚀环境下幼苗库特征的动态变化进行了分析,并对其与土壤水分、土壤养分、降雨量、侵蚀类型等的关系以及不同侵蚀环境下能够成功定居幼苗的主要物种进行了简单讨论,这是我们今后进一步研究黄土丘陵沟壑区幼苗更新能力以及幼苗对地上植被恢复贡献潜力的前提。但黄土丘陵沟壑区本身水分缺乏,植被盖度低,土壤侵蚀严重,影响幼苗存活的环境因子众多,而本研究中未探讨的其他环境因子如光照、温度、地上植被等对幼苗的影响,还有待今后进一步的深入研究,从而探讨影响幼苗存活的关键影响因子。
[1] 邝高明,朱清科,刘中奇,等.黄土丘陵沟壑区微地形对土壤水分及生物量的影响[J].水土保持研究,2012,19(3):74-77.
[2] 秦伟,朱清科,刘中奇,等.黄土丘陵沟壑区退耕地植被自然演替系列及其植物物种多样性特征[J].干旱区研究,2008,25(4):507-513.
[3] 王国梁,刘国彬,刘芳,等.黄土沟壑区植被恢复过程中植物群落组成及结构变化[J].生态学报,2003,23(12):2550-2557.
[4] 吕仕洪,向悟生,李先琨,等.红壤侵蚀区植被恢复研究综述[J].广西植物,2003,23(1):83-89.
[5] 马祥华,焦菊英.黄土丘陵沟壑区退耕地自然植被恢复植被特征及其与土壤环境的关系[J].中国水土保持科学,2005,3(2):15-22.
[6] 朱显谟.迅速全面恢复植被是根除河害之本[J].中国水土保持,1999(10):29-31.
[7] 郑粉莉.子午岭林区植被破坏与恢复对土壤演变的影响[J].水土保持通报,1996,16(5):41-44.
[8] 彭少麟.恢复生态学与退化生态系统的恢复[J].中国科学院院刊,2000,20(3):188-192.
[9] 赵丽娅,李锋瑞.围封沙质草甸土壤种子库与幼苗库的特征[J].西北植物学报,2003,23(10):1725-1730.
[10] Hector G S,Escarre J,Sans F X.Factors that limit the emergence and establish ment of the related aliens Senecio inaequidens and S.pterophorus and the native S.malacitanus in Mediterranean cli mate[J].Canadian Jour nal of Botany,2004,82(9):1346-1355.
[11] 程积民,万惠娥,胡相明.黄土高原草地土壤种子库与草地更新[J].土壤学报,2006,43(4):679-683.
[12] 黄忠良,彭少麟,易俗.影响季风常绿阔叶林幼苗定居的主要因素[J].热带亚热带植物学报,2001,9(2):123-128.
[13] 崔长美,王孝安.黄土高原天然柴松纯林不同坡位幼苗更新特性研究[J].中国农学通报,2011,27(4):48-52.
[14] 康冰,王得祥,崔宏安,等.秦岭山地油松群落更新特征及影响因子[J].应用生态学报,2011,22(7):1659-1667.
[15] 谢佳彦,邓志平.杭州五云山米槠种群幼苗大小结构及空间分布格局研究[J].生态学杂志,2003,22(5):35-39.
[16] Tara A,Daniel F.Seedling establishment and life history trade-offs in alpine plants[J].American Jour nal of Botany,2004,91(7):1147-1153.
[17] Matthew A,Brian C.Seedling establish ment shapes the distribution of shade-adapted foresther bs across a topographical moisture gradient[J].Journal of Ecology,2009,97(5):1037-1049.
[18] Jia Y F,Jiao J Y,Wang N.Relationship bet ween Seedling and Mature Vegetation on the Hilly-gullied Loess Plateau[C]∥2010 inter national conference on combating land degradation in agricultural areas,Xi′an,China,2010:1-5.
[19] 王宁,贾艳峰,白文娟,等.黄土丘陵沟壑区退耕地土壤种子库特征与季节变化[J].草业学报,2009,18(3):42-52.
[20] 焦峰,温仲明,王飞,等.黄土丘陵县域尺度整体景观格局分析[J].水土保持学报,2005,19(2):167-170.
[21] 张振国,黄建成,焦菊英,等.安塞黄土丘陵沟壑区退耕地植物群落土壤抗蚀性分析[J].水土保持研究,2008,15(1):28-31.
[22] 贾燕锋,焦菊英,张振国,等.黄土丘陵沟壑区沟沿线边缘植被特征初步研究[J].中国水土保持科学,2007,5(4):39-43.
[23] 焦菊英,马祥华,白文娟,等.黄土丘陵沟壑区退耕地植物群落与土壤环境因子的对应分析[J].土壤学报,2005,42(5):744-752.
[24] 马克平.生物群落多样性的测度方法:Ⅰ.α多样性的测度方法(上)[J].生物多样性,1994,2(3):162-168.
[25] 焦菊英,张振国,贾燕锋,等.陕北丘陵沟壑区撂荒地自然恢复植被的组成结构与数量分类[J].生态学报,2008,28(7):2981-2997.
[26] 杨涛,王得祥,周金星,等.陕北黄土丘陵沟壑区退耕地植物群落演替规律及物种多样性动态研究[J].西北林学院学报,2009,24(5):10-15.
[27] 朱志诚.黄土高原森林草原的基本特征[J].地理科学,1994,14(2):152-156.
[28] 张瑞,曹华,王云强,等.黄土丘陵沟壑区小流域土壤水分空间变异性及其影响因素[J].水土保持研究,2012,19(5):52-57.
[29] 周萍,刘国彬,侯喜禄.黄土丘陵区侵蚀环境不同坡面及坡位土壤理化特征研究[J].水土保持学报,2008,22(1):7-12.
[30] 田媛,李建贵,潘丽萍.梭梭萌生与初期存活的关键影响因素[J].生态学报,2010,30(18):4898-4904.
[31] 王红梅,谢应忠,陈来祥.黄土高原坡地土壤水分动态特征及影响因素[J].宁夏农学院学报,2004,25(4):62-66.
[32] 王凯博,陈美玲,秦娟.子午岭植被自然演替中植物多样性变化及其与土壤理化性质的关系[J].西北植物学报,2007,27(10):2089-2096.
[33] 陈宇,焦菊英,王宁,等.黄土丘陵区撂荒地不同侵蚀带土壤种子库特征[J].水土保持研究,2012,19(1):1-5.
