鸡粪与化肥不同配比对杨树苗根际土壤酶和微生物量碳、氮变化的影响
2013-09-12井大炜邢尚军
井大炜,邢尚军
(1德州学院,山东德州253023;2山东省林业科学研究院,山东济南250014)
土壤生物学活性主要指土壤微生物和酶活性。土壤微生物是土壤中各种生物化学过程的主要调节者,微生物量碳、氮被认为是土壤活性养分的储存库,是植物生长可利用养分的重要来源[1]。土壤酶是土壤中动植物残体分解、植物根系分泌和土壤微生物代谢的产物,是一类具有生物化学催化活性的特殊物质,参与土壤中许多重要的生物化学过程,如腐殖质的合成与分解,有机化合物、高等植物和微生物残体的分解及其转化等[2]。土壤中的酶和微生物活性高低可以代表土壤中物质代谢的旺盛程度,在一定程度上反映作物对养分吸收利用与生长发育状况等,是土壤肥力的一个重要指标[2-4]。
近年来,许多学者对施用化肥[5-6]、猪圈肥[7]、芝麻饼肥[8]、秸秆还田[9-10]和连年翻压绿肥[11]等处理下微生物量和酶活性的变化进行了深入的研究,而且主要集中在小麦、玉米和烤烟等作物,而对鸡粪有机肥及杨树根际土壤微生物等方面的研究较少。鸡粪是植物生产中优质的有机肥,富含氮、磷、钾等植物所必需的养分,与其他家畜家禽粪便相比养分含量居于首位;与饼肥中氮、磷、钾含量相当,且养分比例变幅小,是优质天然复合肥[12]。利用鸡粪生产制成的有机肥和有机无机复合肥对农作物增产明显。据统计,在西瓜、西兰花、茶叶、红茄、大白菜、杨梅等作物上施用鸡粪有机肥,增产幅度在15% ~25%之间[12]。在果蔗上长期施用鸡粪有机肥,可以改变土壤结构,使板结的土壤松化,增加透气性和保肥性[13]。关于鸡粪有机肥对杨树苗根际土壤酶活性及微生物量碳、氮的影响,尤其对杨树苗在年生长过程内系统的动态变化研究尚未见报道。为此,本试验以山东省济宁市三环化工有限公司生产的鸡粪有机肥为供试原料,开展了鸡粪有机肥与化肥的不同搭配比例对一年生I-107欧美杨在不同生长时期内根际土壤酶活性和土壤微生物量碳、氮的动态变化研究,以期揭示不同施肥处理根际土壤的生物化学过程的变化规律,为探讨施用鸡粪有机肥对杨树苗根际土壤的作用机理提供理论依据,并且为杨树苗的培育和施肥管理提供技术参考。
1 材料与方法
1.1 供试材料
试验设在山东省林业科学研究院试验苗圃,供试土壤为潮土,有机质含量6.83 g/kg,全氮0.52 g/kg,速效氮 27.96 mg/kg,速效磷 26.52 mg/kg,速效钾 79 mg/kg,电导率 24.3 μS/m,pH 8.08。供试鸡粪有机肥为山东省济宁市三环化工有限公司提供,N、P2O5和K2O含量分别为1.22%,1.33%和2.71%,有机质含量24.18%,含水量29.02%。所用化肥为尿素、过磷酸钙、氯化钾。杨树扦插苗品种为I-107欧美杨,接穗长15~16 cm,茎粗2 cm,重量25~27 g。
1.2 试验设计
采用盆栽试验,设5个处理:CK,不施肥;N100,100% 的氮由尿素提供(尿素7.72 g);M10N90,10%的氮由鸡粪有机肥提供(鸡粪有机肥37.54 g),90%的氮由尿素提供(尿素6.95 g);M30N70,30%的氮由鸡粪有机肥提供(鸡粪有机肥112.62 g),70%的氮由尿素提供(尿素5.40 g);M50N50,50%的氮由鸡粪有机肥提供(鸡粪有机肥187.70 g),50%的氮由尿素提供(尿素3.86 g)。每个处理20盆,共计100盆。随机区组排列,除CK外,各处理均为等养分量,N、P和K含量分别为3.55、1.94、3.94 g,各处理P和K不足部分分别用过磷酸钙、氯化钾补足。试验用盆为塑料盆,盆高20 cm、宽30 cm。于2011年4月13日盆栽试验时,将肥料与土壤充分混匀后装盆,每盆装土10.5 kg。
1.3 测定项目和方法
分别在施肥后的30 d、60 d、90 d、120 d、150 d和180 d采集土壤样品,每次每个处理采3盆。在土壤水分含量适中时采用剥落分离法[14]取样,将带土植株取出,先轻轻抖落大块不含根系的土壤,然后用力将根表面附着的土壤全部抖落下来,迅速装入塑料袋内(根际土)。部分新鲜样品立即进行土壤微生物碳、氮的测定,部分土样风干过筛后进行土壤酶活性测定。土壤酶活性和微生物量碳、氮均为每处理3盆土样分别测定。
土壤微生物量碳、氮用氯仿熏蒸法[15-16]测定;土壤脲酶用苯酚钠-次氯酸钠比色法;磷酸酶用磷酸苯二钠比色法;过氧化氢酶用KMnO4滴定法;蔗糖酶用Na2S2O3滴定法[17]测定。
试验数据采用SAS 9.2和Excel 2003软件进行分析。
2 结果与分析
2.1 对杨树苗根际土壤酶活性的影响
2.1.1对根际土壤脲酶活性的影响 不同处理的脲酶活性动态变化规律相似,均呈现先升高后降低的趋势(图1)。在生长前期(30~60 d)时,各处理的脲酶活性缓慢上升,但从生长前期进入快速生长期(90 d)时,各处理均有迅速升高的趋势,并在120 d时达到了最大值,此时M30N70处理的脲酶活性分别比其他处理提高了49.04%、26.49%、12.50%和7.83%,随后逐渐降低。各处理在杨树苗不同生长时期内的总体变化规律为 M30N70>M50N50>M10N90>N100>CK,表明施肥明显增加了根际土壤的脲酶活性。在各施肥处理中,鸡粪配施处理显著高于单施化肥处理,以 M30N70处理最高,比对照提高了24.51%~49.04%,比单施化肥处理提高了14.41%~26.49%。在鸡粪有机肥配施的三个处理中,M10N90与M50N50的脲酶活性均显著低于M30N70。可见,在鸡粪有机肥与化肥的不同配施处理中,随着鸡粪有机肥比例的增加,脲酶活性呈先升高后降低的趋势。因此,并非鸡粪有机肥所占的比例越大,土壤的脲酶活性就越强。
2.1.2对根际土壤碱性磷酸酶活性的影响 各处理的碱性磷酸酶活性均呈先降低后升高再降低的趋势(图2)。所有处理均在120 d时出现峰值,此时杨树苗正处于快速生长期,M30N70处理最高,分别比CK、N100、M10N90和 M50N50提高了 23.18%、20.00%、7.51%和8.77%,随后开始缓慢的下降。

图1 不同处理对根际土壤脲酶活性的影响Fig.1 Effects of different treatments on urease activity in rhizosphere soil
在杨树苗的年生长进程中,各处理碱性磷酸酶活性的变化规律为M30N70>M10N90≈M50N50>N100≈CK。M30N70显著高于其他处理,比 N100提高了13.87% ~27.93%;N100处理与CK差异不显著,可能是由于对照土壤缺磷,土壤供磷能力远不能满足作物生长的需要,因此作物根系分泌较多的磷酸酶,以促进土壤中有机磷化合物水解,生成可以被植物所利用的无机态磷,这是作物适应营养环境的一种机制[18]。配施鸡粪有机肥的处理均显著高于单施化肥处理,说明配施鸡粪可促进土壤碱性磷酸酶活性的提高。另外,在配施鸡粪有机肥处理中,M10N90与M50N50差异不显著,这表明杨树苗根际土壤碱性磷酸酶活性并不随着鸡粪有机肥所占比例的增加而提高,而是取决于鸡粪有机肥与化肥的搭配比例。

图2 不同处理对根际土壤碱性磷酸酶活性的影响Fig.2 Effects of different treatments on alkaline phosphatase activity in rhizosphere soil
2.1.3对根际土壤过氧化氢酶活性的影响 各处理的过氧化氢酶活性均表现出先升高后降低的趋势(图3)。在施肥后120 d,所有处理均达到了最大值,其中,M30N70处理的过氧化氢酶活性最高,分别比 CK、N100、M10N90和 M50N50,提高了 32.37%、21.16%、10.63%和8.53%。之后各处理均开始迅速下降,至落叶期(180 d)时达到了最低值,相比最大值分别降低了 58.96%、49.21%、44.93%、42.79%和44.55%。在杨树苗不同的生长时期内,各处理根际土壤过氧化氢酶活性的变化规律为M30N70>M10N90≈M50N50>N100>CK。M30N70处理分别比CK提高了12.33% ~71.83%,比N100处理提高了12.07% ~27.08%;M10N90处理与M50N50处理差异不显著,但均显著高于CK和N100处理。施肥后80~130 d是杨树苗快速生长、干物质积累最大的时期,也是杨树苗对营养元素吸收量最大的阶段。此时过氧化氢酶活性处于较高值,可以解除杨树苗快速生长积累的过氧化氢等物质产生的毒害,说明过氧化氢酶活性与杨树苗生长发育进程密切相关。

图3 不同处理对根际土壤过氧化氢酶活性的影响Fig.3 Effects of different treatments on catalase activity in rhizosphere soil
2.1.4对根际土壤蔗糖酶活性的影响 所有施肥处理的蔗糖酶活性均显著高于CK,说明施肥能明显提高根际土壤的蔗糖酶活性(图4)。各处理均呈现先升高后降低又缓慢升高的变化趋势。施肥后30~120 d期间,各处理均处于上升的趋势,在120 d时达到最大值,之后开始显著下降,在150 d之后又开始缓慢升高。
在120 d时,M30N70处理分别比 CK、N100、M10N90和 M50N50处理提高了 47.62%、29.17%、9.81%和8.04%;同 N100处理相比,M10N90、M30N70和M50N50处理分别提高了17.63%、29.17%和19.55%。在整个生育期内,M30N70处理的蔗糖酶活性比N100处理提高了29.17% ~46.43%,比CK提高了47.62% ~66.67%。而 M10N90和 M50N50处理分别比 N100处理提高了 14.63% ~27.09%和19.55%~44.33%。说明配施鸡粪有机肥比单施化肥更有利于提高土壤蔗糖酶的活性。在3个配施鸡粪有机肥处理中,随着鸡粪有机肥所占比例的提高,蔗糖酶活性呈现先升高后降低的趋势。
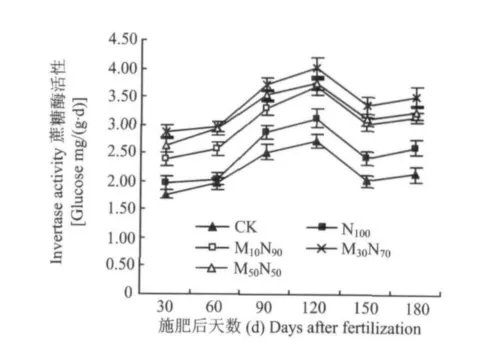
图4 不同处理对根际土壤蔗糖酶活性的Fig.4 Effects of different treatments on invertase activity in rhizosphere soil
2.2 对杨树苗根际土壤微生物量碳、氮的影响
2.2.1对根际土壤微生物量碳的影响 各施肥处理的SMBC含量均显著高于CK(表1)。施肥后60 d,CK达到了最大值,在90 d时就出现了缺肥症状,叶子发黄,根系活性降低,因而土壤微生物活性也降低。而N100、M10N90、M30N70和M50N50处理随着杨树苗的快速生长发育、根系分泌量的增加及气温的不断提高,SMBC含量在增加,到90 d时达到高峰,之后开始迅速地下降。与N100处理相比,M10N90、M30N70和M50N50处理的下降更快。施肥后180 d(落叶期),各处理SMBC含量稍微有所回升,与施肥后150 d相比,分别增加了 27.56、14.14、36.31、35.35和30.73 mg/kg。从杨树苗的年生长进程来看,3个鸡粪有机肥与化肥配施的处理一直显著高于CK和N100处理;M10N90、M30N70和 M50N50处理分别比N100处理提高了47.11~68.39%、63.00% ~76.62%和52.90% ~70.87%。
可见,随着鸡粪有机肥所占比例的增加,SMBC含量呈先增加后减少的趋势。在鸡粪有机肥与化肥的不同配施中,M30N70的SMBC含量最大。结果表明,配施鸡粪有机肥对SMBC含量具有明显的影响,其作用效果取决于鸡粪有机肥与化肥的搭配比例。
2.2.2对根际土壤微生物量氮的影响 在杨树苗的不同生长时期内,总的变化规律为:施肥后30 d各处理土壤SMBN含量达到最高值,随生育进程明显下降,落叶期呈现回升趋势(表2)。施肥后30 d,各施肥处理的SMBN含量均显著高于CK。与N100相比,鸡粪与化肥配施的3个处理均显著提高了SMBN含量。M30N70处理的 SMBN含量显著高于CK、N100、M10N90和 M50N50处理,分别提高了142.17%、82.77%、20.90%和15.11%。施肥后60 d,各处理的SMBN含量均出现不同程度的下降,其中CK的SMBN含量在施肥后90 d最早降为最低值,可能是因为土壤氮素营养不能满足杨树苗生长的需求,土壤微生物不得不加速自身矿化来提供养分,说明在没有氮肥投入的情况下,土壤微生物生物量氮也能够为作物生长提供一定量的氮素养分。而各施肥处理的SMBN含量继续下降,在施肥后150 d达到了最低值;在落叶期(180 d)时,各处理的SMBN含量均有不同程度的升高。

表1 不同处理对根际土壤微生物量碳的影响(mg/kg)Table 1 Effects of different treatments on SMBC in rhizosphere soil

表2 不同处理对根际土壤微生物量氮的影响(mg/kg)Table 2 Effects of different treatments on SMBN in rhizosphere soil
在杨树苗的年生长进程中,鸡粪配施处理均显著地高于对照和单施化肥处理,其中M30N70处理的SMBN含量一直处于最高,分别比CK和N100提高了83.20% ~269.41%和40.01% ~90.38%。可见,在3个鸡粪有机肥与化肥的不同配施处理中,随着鸡粪有机肥比例的增加,SMBN含量呈现先升高后降低的趋势,这与SMBC含量变化规律相似。
3 讨论
3.1 对杨树苗根际土壤酶活性的影响
武雪萍等[8]研究表明,芝麻饼肥和化肥配施与单施化肥相比,能显著提高根际土壤的脂肪酶、转化酶和脲酶活性。李娟等[10]和程东娟等[19]通过田间有机无机肥料定位培肥试验发现,有机无机肥料配施可明显增加土壤脲酶和碱性磷酸酶活性,并且明显高于单施化肥。本研究结果与前人的报道基本一致,鸡粪有机肥与化肥配施明显增强了土壤酶活性,不仅蔗糖酶、脲酶和碱性磷酸酶活性均显著高于对照和单施化肥处理,而且过氧化氢酶活性亦符合这个规律。原因可能是:1)鸡粪有机肥和化肥配施有利于改善土壤理化性质,调节土壤C/N,促进杨树苗和土壤微生物的生长,提高土壤酶活性;2)鸡粪有机肥的施用可以为土壤酶提供更多、更丰富的酶促基质,发挥底物诱导作用;3)施用鸡粪有机肥可以提高土壤腐殖质含量,而腐殖质能够通过离子交换、离子键或共价键等与土壤酶结合,固定土壤酶[20-21]。本试验结果还可看出,在鸡粪有机肥与化肥的不同比例搭配中,比例为3∶7的效果好于1∶9和5∶5的比例。这一方面可能是因为鸡粪有机肥与化肥以3∶7的比例配施更好地调节了土壤的C/N,改善了土壤理化性质,有助于增强根系活性,促进杨树苗的旺盛生长,从而使根系分泌较多的酶类进入土壤;另一方面可能是由于3∶7的比例搭配也促进了土壤动物的生长和微生物的繁殖,加速了有机物的分解,为土壤酶提供了更多的底物。
3.2 对杨树苗根际土壤微生物量碳、氮的影响
许多研究已经证明微生物量是土壤中有机质养分的一种短暂而最有效的贮存形式,是土壤养分的源和库[22]。土壤微生物量作为土壤肥力水平的活指标,已日渐受到土壤科学工作者的关注,研究土壤微生物生物量对了解土壤养分转化及循环具有重要意义[23]。本研究结果表明,在杨树苗的年生长过程中,鸡粪有机肥配施处理的SMBC、SMBN含量均显著高于对照和单施化肥处理,这与 Goyal等[24]和Simek等[25]的研究结果一致。这是由于施肥直接增加根系生物量及根系分泌物,促进微生物生长;同时施用有机肥不但增加了土壤养分,也为微生物提供了充足的碳源,这也证明了鸡粪有机肥配施下有较多的氮素通过同化作用转入到微生物体内被暂时固定,相应地减少了通过NH3挥发和NO-3淋失以及反硝化脱氮等途径造成的氮素损失。这对调节土壤氮素供应、提高土壤氮素利用率、保护大气环境、防止水资源污染和保证林业可持续发展都具有积极意义[26]。
本研究还发现,试验各处理SMBN含量在杨树苗的生育过程中表现为施肥初期升高,随生育进程推进逐渐下降,落叶期有一定回升的趋势。这与罗兰芳等[27]报道一致,这一变化趋势的产生与杨树苗对养分的吸收,土壤微生物活动以及外界气温的变化有关。施肥初期(30 d),各处理SMBN含量达到最高。其主要原因有:1)施入土壤的氮肥迅速释放,土壤矿质氮含量达到最高,而杨树正处于苗期,生长缓慢,对土壤养分的吸收能力很弱;2)随着杨树苗根系的成活,根系分泌物增多,为微生物的活动增加了能源;3)土壤温度的上升,根系周围土壤微生物活性迅速增强,土壤微生物对土壤中氮素开始固持,使土壤微生物生物量增加。随着生育进程的推进,杨树苗对土壤氮素养分的需求量增加,而各处理的SMBN含量逐渐降低,表明一部分微生物量氮又被释放出来,以供杨树苗生长发育的需要,这反映出土壤微生物量氮是植物有效氮的重要储备以及在协调土壤氮素供应方面的重要作用[28]。进入落叶期,杨树苗停止生长,地下根系逐渐衰老死亡,植株通过根系分泌物和残体向土壤提供有机碳、氮,促进了微生物生长,从而使 SMBC、SMBN含量增加[29]。此外,在鸡粪有机肥与化肥的不同比例配施中,3∶7比例搭配处理的SMBC、SMBN含量均显著高于1∶9和5∶5比例搭配处理。可能是因为3∶7比例搭配更好地调节了土壤碳氮比,有利于土壤微生物的大量繁殖和微生物活性的提高,从而增加了土壤微生物生物量。
[1] 王晓龙,胡峰,李辉信,等.红壤小流域不同土地利用方式对土壤微生物量碳氮的影响[J].农业环境科学学报,2006,25(1):143-147.Wang X L,Hu F,Li H X et al.Effects of different land used patterns on soil microbical carbon and nitrogen in small red soil watershed[J].J.Agro-Environ.Sci.,2006,25(1):143-147.
[2] Burns R G.Soil enzymes[M].London:Academic Press,1978.1-33.
[3] Frankenberger W T,Dick W A.Relationship between enzyme activities and microbial growth and activity indices in soil[J].Soil Sci.Soc.Am.J.,1983,47:945-951.
[4] Mijangos I,Perez R,Albizu I et al.Effects of fertilization and tillage soil biological parameters[J]. Enzyme Microbiol.Technol.,2006,40:100-106.
[5] 王继红,刘景双,于君宝,王金达.氮磷肥对黑土玉米农田生态系统土壤微生物量碳、氮的影响[J].水土保持学报,2004,18(1):35-38.Wang J H,Liu J S,Yu J B,Wang J D.Effect of fertilizing N and P on soil microbial biomass carbon and nitrogen of black soil corn agroecosystem[J].J.Soil Water Conserv.,2004,18(1):35-38.
[6] 王光华,齐晓宁,金剑,等.施肥对黑土农田土壤全碳、微生物量碳及土壤酶活性的影响[J].土壤通报,2007,38(4):661-666.Wang G H,Qi X N,Jin J et al.Effect of fertilization on total soil C,microbial biomass C and soil enzyme activities in farmland black soil[J].Chin.J.Soil Sci.,2007,38(4):661-666.
[7] 徐永刚,宇万太,马强,周桦.长期不同施肥制度对潮棕壤微生物生物量碳、氮及细菌群落结构的影响[J].应用生态学报,2010,21(8):2078-2085.Xu Y G,Yu W T,Ma Q,Zhou H.Effects of long-term fertilizations on microbialbiomassC and N and bacterial community structure in an aquic brown soil[J].Chin.J.Appl.Ecol.,2010,21(8):2078-2085.
[8] 武雪萍,刘增俊,赵跃华,等.施用芝麻饼肥对植烟根际土壤酶活性和微生物碳、氮的影响[J].植物营养与肥料学报,2005,11(4):541-546.Wu X P,Liu Z J,Zhao Y H et al.Effects of sesame cake fertilizer on soil enzyme activities and microbial C and N at rhizosphere of tobacco[J].Plant Nutr.Fert.Sci.,2005,11(4):541-546.
[9] 李花,葛玮健,马晓霞,等.小麦-玉米轮作体系长期施肥对土微生物量碳、氮及酶活性的影响[J].植物营养与肥料学报,2011,17(5):1140-1146.Li H,Ge W J,Ma X X et al.Effects of long-term fertilization on carbon and nitrogen and enzyme activities of soil microbial biomass under winter wheat and summer maize rotation system[J].Plant Nutr.Fert.Sci.,2011,17(5):1140-1146.
[10] 李娟,赵秉强,李秀英,So H B.长期不同施肥条件下土壤微生物量及土壤酶活性的季节变化特征[J].植物营养与肥料学报,2009,15(5):1093-1099.Li J,Zhao B Q,Li X Y,So Hwat Bing.Seasonal variation of soil microbial biomass and soil enzyme activities in different longterm fertilizer regimes[J].Plant Nutr.Fert.Sci.,2009,15(5):1093-1099.
[11] 刘国顺,李正,敬海霞,等.连年翻压绿肥对植烟土壤微生物量及酶活性的影响[J].植物营养与肥料学报,2010,16(6):1472-1478.Liu G S,Li Z,Jing H X et al.Effects of consecutive turnover of green manures on soil microbial biomass and enzyme activity[J].Plant Nutr.Fert.Sci.,2010,16(6):1472-1478.
[12] 刘高峰.有机营养对烤烟生理代谢与品质影响的研究[D].福州:福建农林大学硕士学位论文,2006.Liu G F.Studies on the effects of organic nutrients on metabolism,yield and quality of flue-cured tobacco[D].Fuzhou:Ms thesis, Fujian Agricultural and Forestry University,2006.
[13] 赵明,陈雪辉,赵征宇,等.鸡粪等有机肥料的养分释放及对土壤有效铜、锌、铁、锰含量的影响[J].中国生态农业学报,2007,15(2):47-50.Zhao M,Chen X H,Zhao Z Y et al.Releasing of N,P and K of organic fertilizers and their effects on the contents of available Cu,Zn,Fe and Mn in soil[J].Chin.J.Eco-Agric.,2007,15(2):47-50.
[14] Wang X P,Zabowski D.Nutrient composition of Douglas-fir rhizosphere and bulk soil solutions[J].Plant Soil,1998,200:13-20.
[15] 王岩,沈其荣,史瑞和.有机无机肥料施用后土壤生物量C、N、P的变化及N素转化[J].土壤学报,1998,35(2):227-233.Wang Y,Shen Q R,Shi R H.Changes of soil microbial biomass C,N and P and the N transformation after application of organic and inorganic fertilizers[J].Acta Pedol.Sin.,1998,35(2):227-233.
[16] Vance E D.An extraction method form easuring soil microbial biomass C[J].Soil Biol.Biochem.,1987,19(6):703-707.
[17] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.Guan S Y.Soil enzyme and its study method[M].Beijing:Agricultural Press,1986.
[18] 孙瑞莲,赵秉强,朱鲁生,等.长期定位施肥田土壤酶活性的动态变化特征[J].生态环境,2008,17(5):2059-2063.Sun R L,Zhao B Q,Zhu L S et al.Dynamic changes of soil enzyme activities in long-term fertilization soil[J]. Ecol.Environ.,2008,17(5):2059-2063.
[19] 程东娟,刘树庆,王殿武,等.长期定位培肥对土壤酶活性及土壤养分动态变化影响[J].河北农业大学学报,2003,26(3):33-36.Cheng D J,Liu S Q,Wang D W et al.The effect of long-term experiment improving soil fertility on the dynamical changes of soil nutrient and soil enzyme activities[J].J.Agric.Univ.Hebei,2003,26(3):33-36.
[20] Burket J Z,Dick R P.Microbial and soil parameters in relation to N mineralization in soils of diverse genesis under differing management systems[J].Biol.Fert.Soils,1998,27:430-438.
[21] Eivazi F,Bayan M R.Select soil enzyme activities in the historic sanborn field as affected by long-term cropping systems[J].Commun.Soil Sci.Plant Anal.,2003,34:2259-2275.
[22] Ritz K,Wheatley R E.Effects of water amendment on basal and substrate induced respiration rates of mineral soils[J].Biol.Fert.Soils,1989,8:242-246.
[23] Dalal R C,Henderson P A,Glasby J M.Organic matter and microbial biomass in a vertisol after 20yr of zero tillage[J].Soil Biol.Biochem.,1991,23:435-441.
[24] Goyal Sneh C K,Mundra M C,Kapoor K K.Influence of inorganic fertilizers and organic amendments on soil organic matter and soil microbial properties under tropical conditions[J].Biol.Fert.Soils,1999,29:196-200.
[25] Simek M,Hopkins D W,Kaláík J et al.Biological and chemical properties of arable soils affected by long-term organic and inorganic fertilizer applications[J].Biol.Fert.Soils,1999,29:300-308.
[26] 周卫军,曾希柏,张杨珠,等.施肥措施对不同母质发育的稻田生态系统土壤微生物量碳、氮的影响[J].应用生态学报,2007,18(5):1043-1048.Zhou W J,Zeng X B,Zhang Y Z et al.Effects of fertilization on microbial biomass C and N in paddy soils derived from different parent materials[J].Chin.J.Appl.Ecol.,2007,18(5):1043-1048.
[27] 罗兰芳,聂军,郑圣先,等.施用控释氮肥对稻田土壤微生物生物量碳、氮的影响[J].生态学报,2010,30(11):2925-2932.Luo L F,Nie J,Zheng S X et al.Effect of controlled release nitrogen fertilizer on soil microbial biomass carbon and nitrogen contents[J].Acta Ecol.Sin.,2010,30(11):2925-2932.
[28] 何振立.土壤微生物量及其在养分循环和环境质量评价中的意义[J].土壤,1997,(2):61-69.He Z L.Soil microbial biomass and its significance in soil nutrition cycling and soil environment quality assessment[J].Soils,1997,(2):61-69.
[29] Goyal S,Chander K,Mundra M C,Kapoor K K.Influence of inorganic fertilizers and organic amendments on soil organic matter and soil microbial properties under tropical conditions[J].Biol.Fert.Soils,1999,29:196-200.
