不同地理种群茶尺蠖适合度差异研究
2013-09-11王晓庆盛忠雷
王晓庆, 盛忠雷, 彭 萍, 郭 萧, 胡 翔, 林 强
(重庆市农业科学院茶叶研究所,重庆 402160)
茶尺蠖[Ectropis obliqua (Prout)],属鳞翅目尺蛾科(Geometridae),分布遍及各主产茶区,是我国常发性主要茶树害虫之一。为了更好地预防和控制其危害,前人已对安徽郎溪、浙江余杭、江苏无锡等茶尺蠖种群的生物学特性、发生规律以及防治方法等进行了研究[1-8]。不同地理分布区域的同种昆虫,生物学特性因其栖息地气候条件的差异发生相应的变化,导致其发育历期、繁殖力等的地理变异,即昆虫对不同气候环境的适应导致种内的遗传分化,这种现象反映了昆虫生活史对策的多样性[9]。目前,已有报道表明茶尺蠖存在不同的地理宗[10],席羽等人也推测茶尺蠖不同种群的遗传分化与病毒敏感性水平的差异具有潜在的相关性[11]。因此,为明确茶尺蠖种群分化特征与生境适应性的关系,特对不同地理种群茶尺蠖生长发育和生殖力进行研究。
1 材料和方法
1.1 试验材料
试验用的茶尺蠖4个种群分别采自湖北武汉、浙江杭州、江西南昌、福建福安,各采集地的经纬度见表l。采集的幼虫分别于养虫笼(50cm×50cm×60cm)内用水培茶枝饲养,隔日更换茶枝。饲养条件为温度(24±2)℃,相对湿度为70%±5%。实验室饲养1~2代后,收集卵块供本研究使用。
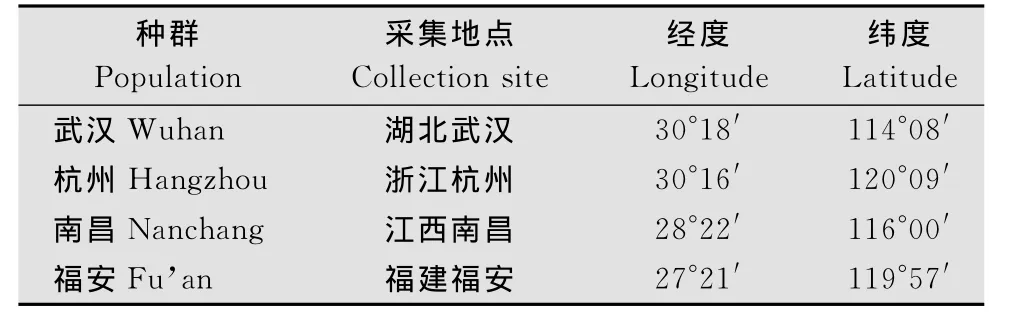
表1 茶尺蠖不同地理种群采集地信息Table 1 Sampling site information of E.obliqua
1.2 试验方法
试验设4个地理种群,每个种群设置4个重复。处理自卵开始,放入置有滤纸的培养皿内保湿,每日记录其孵化幼虫数。幼虫自孵化后即1龄开始,饲以茶树鲜叶,每日更换新鲜叶片并清理虫粪,直到幼虫停止取食。幼虫老熟后,在瓶内放入含水量10%~15%的消毒土壤供其化蛹。成虫羽化配对后,放入体积为2L的透明塑料罩,顶部用纱布封口,饲以10%蜂蜜水,罩内侧放长约10cm的折叠绘图纸供产卵用。每个重复30头虫,每处理连续饲养3个世代(依次命名为F1、F2和F3代)。逐日检查茶尺蠖发育状况、死亡数、产卵量等种群参数,各处理均在人工气候箱内,温度为(25±1)℃,相对湿度为70%±5%,光周期为L∥D=16h∥8h。
1.3 数据处理
用DPS数据处理系统对试验数据进行统计分析[12],计算均值及标准误,均值差异显著性通过Duncan氏新复极差法进行多重比较。不同地理种群茶尺蠖生命表的组建采用Brich方法[13],相关参数的计算参照徐汝梅介绍的方法[14]。
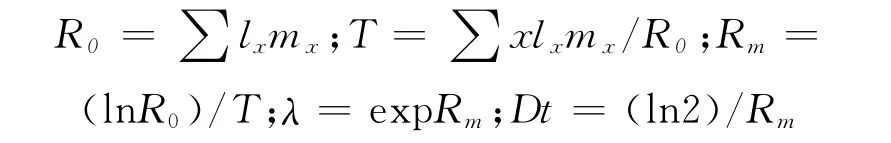
式中:R0为净生殖率,T为平均世代周期,Rm为内禀增长率,λ为周限增长率,Dt为种群加倍时间,x为时间间距,lx为x年龄段的雌虫存活率,mx为x年龄段每雌产雌数。
2 结果与分析
2.1 不同地理种群茶尺蠖(F1代)发育的差异
在试验条件下,不同地理种群茶尺蠖卵和幼虫的发育历期具有显著差异,而蛹和成虫的发育历期基本无差异(表2)。卵发育历期以福安种群最长,南昌种群次之,两者显著长于武汉和杭州两个种群(P<0.05)。在各种群之间,幼虫历期长短表现出显著不同,其中2龄、4龄和预蛹的发育历期无显著差异(P>0.05),1龄幼虫发育历期福安种群最长,南昌种群次之,两者之间达到显著性差异(P<0.05),且显著长于其他两个地理种群(P<0.05);3龄和5龄则以南昌和福安两个种群发育历期长,与武汉和杭州两个种群达到显著性差异(P<0.05)。
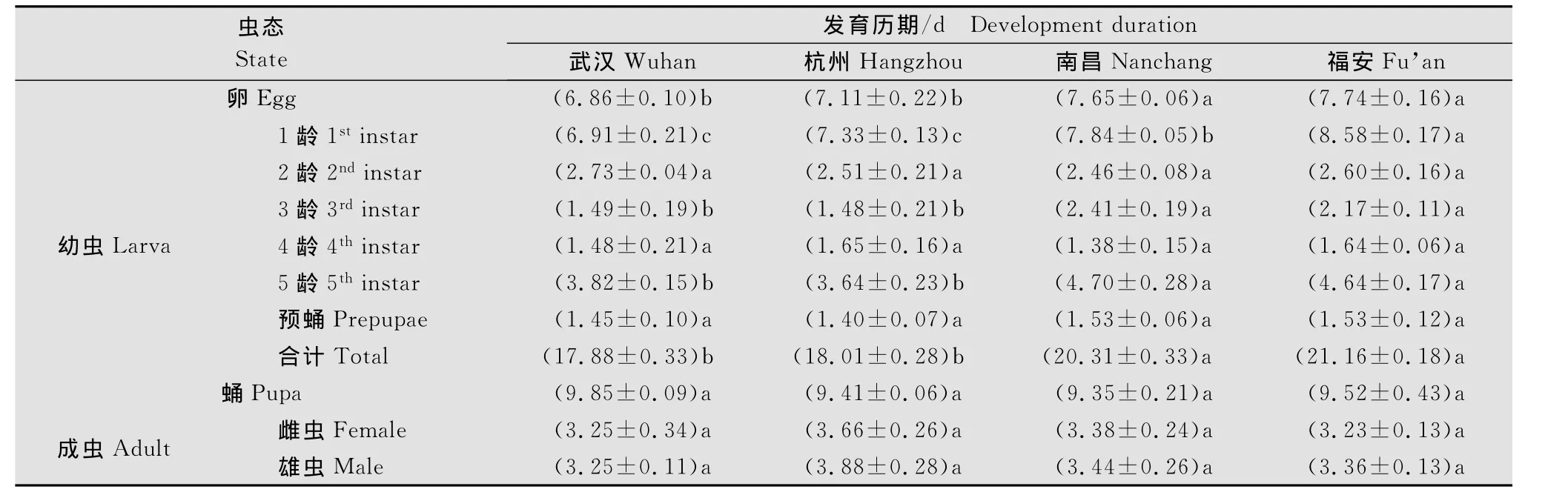
表2 不同地理种群茶尺蠖的发育历期1)Table 2 Development duration of different geographic populations of E.obliqua
2.2 不同地理种群茶尺蠖繁殖力的差异
根据茶尺蠖不同地理种群的生长发育及繁殖结果,组建了茶尺蠖实验种群生命表(表3)。茶尺蠖武汉种群净生殖率(R0)最大,为200.76;世代周期(T)以福安种群最大,说明福安种群发育速率最慢;内禀增长率(Rm)则以武汉种群最大,为0.1447,福安种群最小,为0.1216;周限增长率(λ)的大小排序为武汉>杭州>南昌>福安,而种群加倍时间(Dt)大小正好相反。
方差分析结果表明,4个地理种群中武汉与南昌茶尺蠖的净生殖率无显著性差异(P>0.05),但与杭州和福安间差异显著(P<0.05),同时,杭州、南昌和福安3个种群间无显著性(P>0.05);武汉与杭州,南昌和福安间茶尺蠖的平均世代周期(T)差异不显著(P>0.05),但南昌、福安与武汉、杭州间差异显著(P<0.05);武汉与杭州、南昌和福安种群间茶尺蠖的内禀增长率(Rm)、周限增长率(λ)和种群加倍时间(Dt)均具有显著性差异(P<0.05),而其他三个种群间无明显差异(P>0.05)。
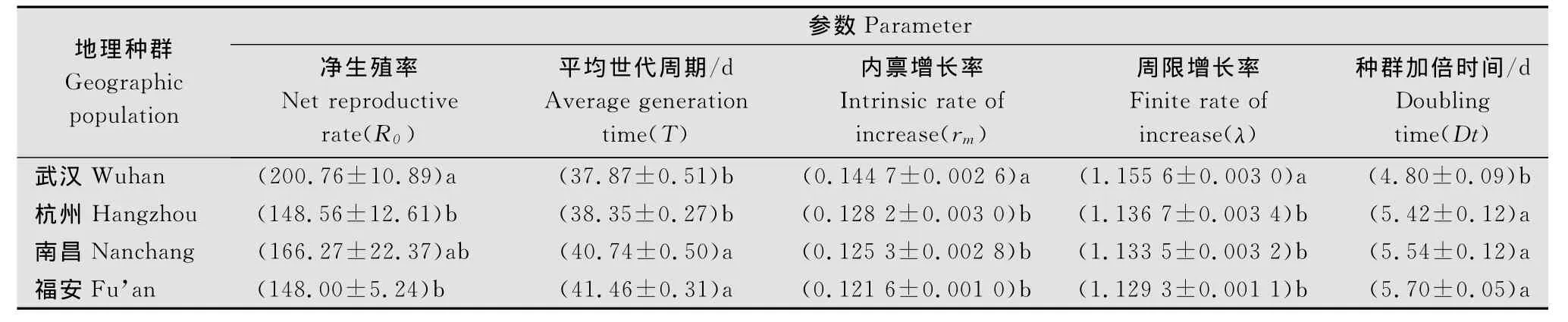
表3 不同地理种群茶尺蠖生命表参数1)Table 3 Life table parameters of different geographic populations of E.obliqua
2.3 不同地理种群茶尺蠖3个世代生长发育的差异
由图1和图2可看出,同一种群,不同世代茶尺蠖间的幼虫发育历期和平均世代周期随着世代的推移有增加的趋势,但差异不显著(P>0.05)。同时,在室内连续饲养茶尺蠖3个世代中,均属福安的幼虫发育历期和平均世代周期最长,武汉的最短。不同地理种群和不同世代间平均世代周期变化趋势和幼虫发育历期基本一致。

图1 不同地理种群的不同世代茶尺蠖幼虫发育历期Fig.1 The larval developmental duration of different geographic populations of E.obliqua
2.4 不同地理种群茶尺蠖3个世代存活率和生殖力的差异
图3分析结果显示,不同地理种群茶尺蠖的存活率F1代与F2和F3代间差异显著(P<0.05),且F2代与F3代间差异不显著(P>0.05)。即室内连续饲养茶尺蠖,F2代存活率显著下降,但F3代与F2代相比,存活率虽有所下降,而差异不明显,基本趋于稳定。

图2 不同地理种群的不同世代茶尺蠖平均世代周期Fig.2 Average generation time of different geographic populations of E.obliqua
同一世代茶尺蠖不同地理种群间存活率均属武汉种群最高,福安最低(图3)。在F1代和F2代中,武汉种群茶尺蠖的存活率显著高于福安种群(P<0.05),但与杭州和南昌两个种群间无显著差异(P>0.05),且后3个种群间也无显著性差异(P>0.05);在F3代,武汉种群的存活率显著高于杭州、南昌和福安3个种群(P<0.05),但后3个种群间无显著性差异(P>0.05)。
不同地理种群茶尺蠖的平均每雌产卵量世代间差异较大(图4)。4个地理种群的平均每雌产卵量随着世代的推移均有所下降,其中武汉和福安两个种群F1代与F2代间均未达到显著性差异(P>0.05),但与F3代间差异显著(P<0.05);南昌种群F1代与F2代和F3代间差异显著(P<0.05),但杭州种群世代间差异不显著(P>0.05)。
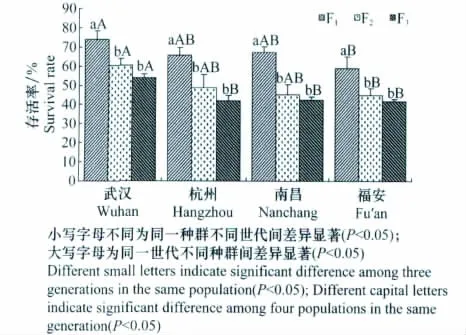
图3 不同地理种群的不同世代茶尺蠖世代存活率Fig.3 The survival rate per generation of different geographic populations of E.obliqua
同一世代茶尺蠖不同地理种群间的平均每雌产卵量均无显著性差异(P>0.05)(图4)。在F1代,南昌种群茶尺蠖的平均每雌产卵量稍高于其他种群,但在F2和F3代,各种群间的产卵量相差不大。
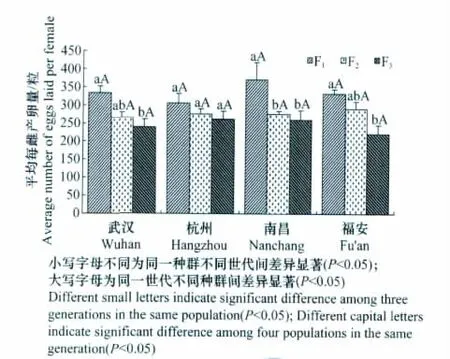
图4 不同地理种群的不同世代茶尺蠖平均每雌产卵量Fig.4 The average number of eggs laid per female of different geographic populations of E.obliqua
3 讨论
资料表明,在相同温度下,同种昆虫高纬度和低纬度种群发育历期长短因种群不同而异。本研究结果表明,纬度偏低地区的茶尺蠖种群卵和幼虫的发育历期长于纬度稍高地区的种群,即在同一室内条件下,福安种群发育慢,武汉种群发育快。而处于相似纬度的安徽郎溪[15]和浙江杭州[16]地理种群,其幼虫发育历期也存在差异。这表明即使在同一类昆虫中,其发育历期的地理变异与纬度和微域气候均有关系。同时,本文结果得出蛹和成虫种群间发育历期差异不明显,揭示了茶尺蠖生长发育存在地理变异现象,但变异程度因虫态不同而异。
本试验选用的4个地理种群,在相同饲养条件下,武汉种群茶尺蠖的内禀增长率及周限增长率最大,种群的加倍时间最短。这表明武汉种群因环境的选择压力而使其生物学特性出现了明显的差异。虽然实验种群生命表所表明的种群动态和自然种群的真实反映有很大差别,但它却为研究自然种群的动态以及计算机模拟奠定了基础[17]。
本研究发现,地理种群间茶尺蠖发育历期的差异不会随试验条件下饲养的时间延长而减小,这与之前的研究棉铃虫安阳种群和四川种群放在室内同样条件下饲养3个世代,第一代各虫态发育差异较大,后两代差异显著缩小的现象相异[18]。而本研究中茶尺蠖种群间存活率和生殖力随着试验条件下不断的饲养,差异会逐渐减小。
[1] 高旭晖,宛晓春,杨云秋,等.茶尺蠖生物学习性研究[J].植物保护,2007,33(3):110-113.
[2] 林少和.茶尺蠖的发生规律及防治方法[J].福建农业科技,2003(1):52-53.
[3] 殷坤山,熊兴平,单夏锋.茶尺蠖发育历期和有效积温的研究[J].植物保护,1995,21(1):16-18.
[4] 楼云芬.茶尺蠖发育历期的温度效应[J].茶叶科学,1993,13(2):127-133.
[5] 彭萍,王晓庆,邓敏,等.20%氯虫苯甲酰胺防治茶尺蠖药效试验[J].植物医生,2010,23(3):23-24.
[6] 刘琴.茶尺蠖病毒与苏云金杆菌增强防治茶尺蠖作用效果的研究和应用[D].南京:南京农业大学,2005.
[7] 陈宗懋,许宁,韩宝瑜,等.茶树-害虫-天敌间的化学信息联系[J].茶叶科学,2003,23(S1):38-45.
[8] 胡萃,朱俊庆,叶恭银,等.茶尺蠖[M].上海:上海科学技术出版社,1994:28-31.
[9] 涂小云.亚洲玉米螟滞育和生物学特性地理变异的研究[D].南昌:江西农业大学,2011.
[10]郑加锋.茶尺蠖三个地理宗[J].福建茶叶,1988(2):38.
[11]席羽.茶尺蠖地理种群对茶尺蠖核型多角体病毒的敏感性差异及遗传变异研究[D].北京:中国农业科学院,2011.
[12]唐启义,冯明光.实用统计分析及其DPS数据处理系统[M].北京:科学出版社,2002:53-55.
[13]Birch L C.The intrinsic rate of natural increase of an insect population[J].The Journal of Animal Ecology,1948,117:15-26.
[14]徐汝梅.昆虫种群生态学[M].北京:北京师范大学出版社,1987:97-107.
[15]张汉鹄.茶尺蠖发育起点温度与有效积温研究[J].安徽农业科学,1989(1):62-66.
[16]朱骏庆.茶尺蠖发育起点与有效积温的研究[J].茶业通报,1990(1):32-34.
[17]王进军,赵志模,李隆术.嗜卷书虱实验种群生命表的研究[J].应用生态学报,2001,12(1):83-85.
[18]李锦华.不同地理种群棉铃虫饲养比较[J].昆虫知识,1982,21(4):7-8.
