番茄温敏型核雄性不育系T-4花药蛋白质含量、POD及EST同工酶的研究
2013-09-03国亚慧于分弟甘桂云韦冠睦王先裕杜永臣桝田正治
国亚慧 于分弟 甘桂云 韦冠睦 王先裕,6* 杜永臣 桝田正治
(1广西大学农学院,广西南宁 530004;2广西来宾合山市岭南镇人民政府,广西来宾 546500;3广西柳州市农业科学研究所,广西柳州 545003;4中国农业科学院蔬菜花卉研究所,北京100081;5日本冈山大学,日本700-8530;6国家大宗蔬菜产业技术体系桂林综合试验站,广西桂林 541004 )
番茄(Lycopersicon esculentumMill.)属于茄科番茄属(Lycopersicon),自花授粉作物。植物育性是由基因控制的生理过程、生化反应和形态构建的最终结果。物质代谢是植物生命活动的基础,是植株间的生理生化差异的反映。在发育过程中正常发育的花药和花粉会积累一些物质,如蛋白质、核酸、碳水化合物(可溶性糖和淀粉)和某些氨基酸等,而雄性不育系花药或花粉则相对缺乏。夏涛和刘纪麟(1993)在研究玉米细胞质雄性不育系的物质代谢时,提出了“物质亏损”的概念,提出结构物质、营养物质、能量物质的亏损现象严重影响小孢子的发育。国内外已对水稻(孙谷畴 等,1997;张明永 等,1997)、油菜(蒋梁材和刘启鑫,1994)、小麦(张爱民 等,1996;张花 等,1996)、大白菜(王淑华 等,1998)、辣椒(耿三省 等,1997)及甜菜(孙立险,1986)、榨菜(胡美华 等,2000)等作物的不育花粉(药)或叶片中氨基酸、同工酶、蛋白质等生化物质的异常进行了研究,揭示了雄性不育的生化机理。
王保仁等(1980)、刘忠松(1987)发现湘矮A CMS(属Pol CMS 胞质)不育系及相应保持系间过氧化物酶(POD)同工酶谱带存在差异;陈松等(1995)研究认为油菜不育系及其同型保持系的EST同工酶谱带也有差异,胞质基因的不同会导致基因产物酯酶(EST)的不同;林宝刚等(2005)对Pol CMS、陕2A CMS及其同一保持系陕2B 的种子进行EST、POD同工酶比较分析后发现,不育材料种子的EST、POD同工酶都与保持系具有较大差异。李殿荣(1986)报道,杂交油菜秦油2号及其亲本植株不同营养器官的EST、POD同工酶酶谱间存在着差异。张子学等(2008)研究表明,辣椒不育系与保持系材料的POD同工酶活性及种类、EST同工酶酶带在花药不同发育时期存在差异,超氧化物歧化酶(SOD)同工酶活性及种类无明显差异。截至目前对番茄不育系同工酶的研究报道甚少。雄性不育系在番茄育种中扮演着重要的角色,不但省工、省力,少去了人工去雄的步骤,还能提高种子质量,减少假种子的混杂。本试验主要对番茄温敏型核雄性不育系T-4的蛋白质、POD及EST同工酶进行研究,探索该材料不育的机理,为更好地研究和利用不育系提供一定的理论基础。
1 材料与方法
1.1 试验材料
试验材料为温敏型核雄性不育系T-4和对照First,是由日本冈山大学农学部作物繁殖研究室桝田正治教授提供。在日本,T-4为对照First经γ射线辐照后所得的变异突变体,当温度达到某一范围时,T-4可恢复部分育性,高于或低于该临界温度范围则表现为雄性不育,临界温度为20 ℃(Masaharu MASUDA and Christopher O.OJIEWO.2006)。不同地区T-4核雄性不育性表现出不同的临界温度,即临界温度迁移现象。在广西南宁地区,夜温为20 ℃左右是T-4育性转换的临界温度,夜温高于临界温度时表现为不育,低于14 ℃时也表现为不育(王先裕 等,2011)。
1.2 试验方法
材料于2010年8月在广西大学玻璃温室中育苗,两片真叶时转入人工培养箱中进行处理。共设3个处理,处理的温度分别为昼28 ℃/夜24 ℃,昼28 ℃/夜18 ℃以及昼28 ℃/夜12 ℃,每天光照12 h(7:00~19:00),湿度为70%。每个处理4株,对照为First,3次重复。
采集当天开放的花朵,在-80 ℃下保存,用于蛋白质、POD及EST同工酶的测定分析。蛋白质用考马斯亮蓝G-250染色分光光度法测定;POD及EST同工酶采用聚丙烯酰胺凝胶垂直板电泳法进行测定。
蛋白质提取测定:准确称取约0.1 g番茄花药,放入研钵中,加入2 mL蒸馏水在冰浴中研成匀浆,离心(4 000 r·min-1,10 min),将上清液倒入10 mL容量瓶,再向残渣中加入2 mL蒸馏水,悬浮后再离心10 min,合并上清液,定容至刻度。
另取1支具塞试管,准确加入0.1 mL样品提取液,再加入0.9 mL蒸馏水、5 mL考马斯亮蓝G-250试剂,充分混合,放置2 min后,以标准曲线1号试管做参比,在595 nm波长下比色,记录吸光度。根据所测样品提取液的吸光度,在标准曲线上查得相应的蛋白质含量,计算样品蛋白质含量。

花药POD液、EST液的提取:称量0.1 g花药,放入研钵中,加入pH 8.3Tris-HCl和20%的蔗糖缓冲液,在低温下研磨至匀浆,在4 000 r·min-1离心15 min。取上清液放入超低温恒温冰箱-20 ℃备用。
染色方法:POD同工酶采用联苯胺染色法;EST同工酶用α-醋酸萘酯、β-醋酸萘酯及坚牢蓝RR盐染色液染色。计算相对迁移率(Rf)。
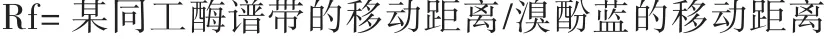
2 结果与分析
2.1 不同温度处理下T-4与对照First花药中蛋白质含量
从表1中可以看出,对照First在3个温度处理下(夜温分别为24 ℃、18 ℃及12 ℃时)的蛋白质含量分别为2.85、2.75 μg·g-1及2.84 μg·g-1,三者之间差异不显著。T-4在昼28℃/夜24 ℃及昼28 ℃/夜12 ℃的处理下的蛋白质含量分别为1.67μg·g-1和1.43μg·g-1,显著低于昼28℃/夜18℃处理;在昼28 ℃/夜18 ℃处理下T-4蛋白质含量与对照First差异不显著。即T-4在不育温度条件下(夜温高于20℃或低于14℃)的蛋白质含量显著低于可育温度条件下的蛋白质含量,且显著低于对照First。

表1 不同温度处理下T-4与对照First花药中蛋白质含量
2.2 不同温度处理下T-4与对照First花药POD及EST同工酶酶谱
从图1可以看出,在3个温度处理下对照First的POD同工酶酶谱无明显差异。T-4在昼28℃/夜18 ℃的可育条件下的POD同工酶酶谱与对照First间无明显差异。T-4在不育温度条件下,即昼28 ℃/夜24℃和昼28 ℃/夜12℃时POD同工酶酶谱,较可育温度条件下多了一条迁移率为0.143的谱带P1。表明T-4在昼28 ℃/夜24℃和昼28 ℃/夜12℃不育温度条件下花药的POD同工酶谱带与可育温度处理存在差异。
从图2可以看出,在昼28℃/夜18℃的可育条件下,T-4 的EST同工酶酶谱共出现了4条酶带;在昼28℃/夜24℃和昼28℃/夜12℃不育条件下,出现6条酶带。即可育温度条件下的EST同工酶酶谱比不育温度条件下少了迁移率分别为0.415及0.491的2条谱带,分别以E3和E4表示。对照First在3个温度处理下酶谱无明显差异。可育温度条件下T-4的EST同工酶酶谱与对照First无显著差异。表明对温敏型雄性不育系T-4进行不同温度处理,EST同工酶的谱带存在差异,可育温度条件下同工酶的谱带少于不育温度处理。
综上所述,在不同温度处理下,T-4花药中POD、EST同工酶酶谱有差异,不育温度条件下的POD、EST同工酶均表现为有较多谱带,而可育温度条件下的谱带则较少。

图1 3个不同温度处理下T-4与对照First花药中POD同工酶酶谱

图2 3个不同温度处理下T-4与对照First花药中EST同工酶酶谱
3 结论与讨论
本试验结果表明,在不育温度条件下T-4的花药中蛋白质含量显著低于可育温度条件下的蛋白质含量,且显著低于对照First花药中蛋白质含量;同工酶酶谱的分析显示,T-4在不育温度条件下花药中的POD同工酶和EST同工酶谱带数量均多于可育温度处理。从试验结果可推测,番茄花药中蛋白质的含量高低及POD同工酶和EST同工酶的表达均对育性表现存在一定的影响。本试验仅从花药中蛋白质的角度对番茄育性表现进行了研究,为了更好地了解番茄雄性不育的发生机理,有待进行全方位地剖析,从而更好地利用番茄雄性不育系在育种中的优势。探索简易的操作方式来对番茄雄性不育系进行筛选是一大难题,目前仅仅从外观形态和简单的花粉萌发情况对雄性不育系的育性进行判定,存在很大的不稳定性和不准确性,为了进一步探索不育系的育性作用机理,本试验从蛋白质水平上对其进行分析,有待从基因水平上对其进行定位。
陈松,蒲惠明,戚存扣.1995.甘蓝型双低油菜三系及杂种酯酶同工酶分析.江苏农业科学,(2):27-28.
耿三省,毛爱军,蒋健箴.1997.辣椒雄性不育花药的生化特性.北京农业科学,15(2):26-27.
胡美华,陈竹君,汪炳良.2000.榨菜胞质雄性不育系及其保持系过氧化物酶和酯酶同工酶的比较研究.浙江农业学报,12(4):201-205.
蒋梁材,刘启鑫.1994.甘蓝型油菜雄性不育系与可育系花蕾的生理生化特性.中国油料,16(1):11-14.
李殿荣.1986.甘蓝型油菜雄性不育系、保持系、恢复系选育成功并大面积推广.中国农业科学,4(4):94.
林宝刚,张明龙,王桂荣.2005.辐照诱导的新雄性不育系过氧化物酶和酯酶同工酶分析.核农学报,19(4):304-306.
刘忠松.1987.高等植物雄性不育的生理生化研究进展.植物生理学通讯,23(2):16-21.
孙谷畴,林植芳,林桂珠,梁承邺.1997.细胞质雄性不育水稻幼穗和花药发育期的蛋白酶活性变化.热带亚热带植物学报,5(4):45-51.
孙立险.1986.甜菜雄性不育系及保持系的过氧化物酶同工酶初步分析.中国糖料,(4):21-25.
王保仁,常桂菊,胡能书.1980.湘矮型油菜“三系”及其杂种过氧化物酶同工酶的初步研究.湖南农业科技,(6):16-21.
王淑华,魏毓棠,冯辉,佟景春.1998.大白菜雄性不育株与可育株花蕾生理生化特性比较分析.沈阳农业大学学报,(2):132-137.
王先裕,于分弟,梁聪耀,李良韶,黄维娜,杜永臣,刘政国,曾东,潘立凡.2011.温敏型核雄性不育系番茄T4育性的研究.中国蔬菜,(14):64-68.
夏涛,刘纪麟.1993.玉米细胞质雄性不育系物质代谢系统的研究.华中农业大学学报,12(1):1-6.
张爱民,李英贤,黄铁城.1996.小麦雄性不育与内源激素关系的初步研究.农业生物技术学报,4(1):56-61.
张花,何之常,徐乃瑜.1996.不同细胞质雄性不育小麦阳离子过氧化物酶和IAA-氧化酶活性的研究.武汉大学学报,42(6):728-732.
张明永,田长恩,梁承邺,黄毓文,刘鸿先.1997.细胞质雄性不育水稻幼穗和花药的呼吸酶活性研究.热带亚热带植物学报,5(4):52-55.
张子学,王丽,王志杰.2008.辣椒雄性不育系与保持系花药POD、SOD和EST同工酶表达差异研究.安徽生物学报,17(4):616-622.
