南方红壤区3种典型森林恢复方式对植物群落多样性的影响
2013-08-02欧阳志云陈法霖陈圣宾
王 芸,欧阳志云,郑 华,陈法霖,陈圣宾,曾 静
(中国科学院生态环境研究中心城市与区域生态国家重点实验室,北京 100085)
森林生态系统具有重要的生态系统服务功能[1-2],对维持我国自然生态系统格局、功能和过程具有重要意义[3]。我国72%的林地处于退化状态[4],生态服务较低,因而,恢复和重建退化森林生态系统是改善我国环境问题的根本所在[5]。合理的森林恢复方式能够提高植物群落的多样性[4,6-7],进而提高生态系统服务功能[8]。因此,研究不同森林恢复方式尤其是未经强烈人工干扰的植物群落构成及多样性[9]对生态恢复重建[10]和生态系统服务功能[2]具有重要意义。
按照是否有人为活动干扰,森林恢复方式有自然恢复和人工恢复,而人工造林是人工恢复的重要内容。本地种马尾松(Pinus massoniana)适应能力强,是我国分布最广的松科植物[11],马尾松人工林的面积占我国亚热带地区的造林面积的14%[12]。引进种湿地松(Pinus elliottii)具有抗旱耐贫瘠、早期生长快等特点[13-14],在我国大面积种植。纯林或者外来种人工林会降低植物群落的多样性[9],改变植物原初的植物群落结构,最终造成森林生态系统服务功能的降低[6,8,14]。自然恢复能够加速森林恢复,具有更高的生态系统服务功能[6,8,15-16]。有Meta分析认为,人工恢复生物多样性、生态系统服务功能高于退化区,而低于原始状态的对照区[7]。当前关于生态恢复对植物群落影响的研究,多集中于不同演替、恢复阶段[10,16],而关于恢复方式对植群落[4,17]乃至生态系统服务功能影响[8]的研究则较少。
从植物群落多样性、土壤肥力维持等角度进行研究,有助于阐明不同森林恢复方式之间生态系统服务功能的差异,为我国南方红壤区的森林恢复提供理论依据,本课题将对相关问题开展系列研究。本文主要从植物群落多样性开展研究,目的在于回答以下两个科学问题:一是南方红壤区自然恢复(天然次生林)和人工恢复(马尾松人工林和湿地松人工林)的植物群落结构特征和多样性有何差异。二是本地种马尾松人工林和引进种湿地人工松人工林植物群落结构特征和多样性有何差异。
1 材料与方法
1.1 研究区自然概况
本研究在湖南省衡阳县、江西省安福县、广西省桂林市的雁山区、临桂县和阳朔县开展,样地地处24.890—27.318°N,110.150—114.549°E之间。样地属亚热带季风湿润气候,年均温17.9—18.9°C,年降雨1452—1950 mm。样地海拔90—295 m,土壤为红壤。研究区的天然次生林自恢复之后,人为干扰较少。马尾松人工林和湿地松人工林在最初种植的3—4 a后抚育1次,包括乔木下表土疏松以及林下植被的砍伐,之后不再进行抚育。所有样地恢复时间约为20年,样地其余详细信息参见文献[18]。
1.2 样地设置与植物群落调查
每种森林恢复方式选取15个独立的样地,每个样地上调查2—3个10 m×10 m面积上所有乔木的株高、胸径和株数,每个乔木样方内调查1—2个5 m×5 m面积上的灌木物种数及株数,1—3个1 m×1 m面积上的草本物种数及株数。共调查乔木样方127个(其中湿地松人工林样方42个,马尾松人工林样方45个,天然次生林样方40个),灌木样方238个(湿地松人工林样方80个,马尾松人工林样方86个,天然次生林样方72个),草本样方357个(其中湿地松人工林样方120个,马尾松人工林样方 129个,天然次生林样方108个),其中乔灌草的分层根据[19]划分。
1.3 数据分析
1.3.1 植物种重要值计算方法[20]
乔木层采用(1)式;灌木和草本层采用(2)式:

式中,Di、Fi和Ai分别代表第i个物种的相对多度、相对频度和相对优势度。相对多度等于第i个物种的个体数除以群落中所有物种的个体数的和;相对频度等于第i个物种在统计样方中出现的次数除以所有种出现的总频数;相对优势度,以相对胸径面积表示,等于第i个物种的胸高断面积除以群落中所有物种的胸高断面积和。
1.3.2 植物区系构成分析
种子植物的植物区系分析参考吴征镒先生的种子植物分布类型进行[21]。
1.3.3 群落多样性指数
物种丰富度即为植物种数。
辛普森指数 Simpson index[22],计算公式如下:

式中,ni指样地中第i个植物种的株数,N指样地中所有植物种总株数。
1.4 数据处理
群落中的优势科按累积重要值确定[23],通过单因素方差法检验恢复方式对植物群落多样性指数的影响。采用去趋势对应分析(DCA)分析不同森林恢复方式植物群落结构差异。植物群落的多元方差分析通过R语言中Vegan数据包的mrpp,adonis和anosim程序实现[24]。单因素方差分析均通过SPSS16.0软件实现;DCA用Canonco 4.5进行;用Sigmaplot软件作图。
2 结果与分析
2.1 植物区系基本构成和优势植物构成
植物的区系构成能反映地理和生态等信息,是研究植物群落多样性的基础。天然次生林有226种植物,隶属86科160属,湿地松人工林有155种,隶属66科118属,马尾松人工林有137种植物,隶属59科97属(表1)。
在乔木层中,湿地松人工林有32种植物,隶属5科6属,马尾松人工林有32种植物,隶属10科11属,天然次生林有121种植物,隶属37科47属(表1)。湿地松人工林中重要值最大的科是松科(Pinaceae),只包含湿地松一个物种,其重要值达到93.0%。马尾松人工林中重要值最大的科是松科,只包含马尾松1个种,其重要值达到 86.1%。天然次生林中重要值较大的科是壳斗科(Fagaceae)、樟科(Lauraceae)和山茶科(Theaceae),其累积重要值达到49.2%。
在灌木层,天然次生林的植物科数、种数和属数高于两种人工林(表1)。湿地松人工林中重要值较大的科按重要值由大到小依次是马鞭草科(Verbenaceae)、禾本科(Gramineae)、金缕梅科(Hamamelidaceae)、山茶科、大戟科(Euphorbiaceae),累积重要值达52.0%;马尾松人工林中重要值最大的科按重要值由大到小依次是金缕梅科、马鞭草科、山茶科、壳斗科和桃金娘科(Myrtaceae),累积重要值达53.2%;天然次生林中重要值较大的科按重要值由大到小依次是壳斗科、金缕梅科、山茶科、蔷薇科(Rosaceae)、茜草科(Rubiaceae)和桃金娘科,累积重要值达49.0%。
在草本层,植物的科数、属数和种数表现为天然次生林高于湿地松人工林和马尾松人工林(表1)。湿地松人工林中重要值较大的科按重要值由大到小依次是禾本科、里白科(Gleicheniaceae)和金星蕨科(Thelypteridaceae),其累积重要值为48.0%。马尾松人工林中重要值较大的科是里白科、金缕梅科和鳞毛蕨科(Dryopteridaceae),累积重要值为50.7%。天然次生林中紫金牛科(Myrsinaceae)、禾本科、鳞毛蕨科、纸莎草科(Cyperaceae)和茜草科重要值较大,累积重要值为50.0%。

表1 3种森林恢复方式的植物群落区系基本构成Table 1 Floral basic composition under 3 forest restoration approaches
2.2 植物区系成分分析
所调查到的植物区系成分以热带成分为主,温带成分次之,世界分布成分最少,分别占总属数的42%—70%、20%—48%和3%—16%(图1)。在湿地松人工林中检测到的区系成分及变型有18种,马尾松人工林有16种,天然次生林有21种。湿地松人工林特有的植物区系成分为热带亚洲、非洲(或东非、马达加斯加)和大洋洲间断分布,天然次生林特有的植物区系成分为热带亚洲、大洋洲(至新西兰)和中、南美(或墨西哥)间断分布、热带亚洲、非洲和中、南美洲间断分布和旧世界温带分布。湿地松人工林比例超过10%的地理区系成分为北温带分布和泛热带分布区系,分别占总属数的34.9%和28.4%。马尾松人工林比例超过10%的地理区系成分为北温带分布、热带亚洲至热带大洋洲分布和泛热带分布,分别占总属数的43.8%、19.2%和18.5%。天然次生林比例最大的区系成分为泛热带分布,占27.8%,其次为热带亚洲至热带大洋洲分布、北温带分布和东亚和北美洲间断分布,分别占14.7%、13.7%和12.4%(图1)。
2.3 植物群落多样性比较
天然次生林每个10 m×10 m的小样方上有(19.2±1.1)种植物,显著高于马尾松人工林14.8±1.3和湿地松人工林14.4±0.7,两种人工林之间的物种丰富度差异不显著(P>0.05)。天然次生林的香农多样性指数显著高于两种人工林,两种人工林之间差异不显著(P>0.05,表2)。
乔木层的物种丰富度和多样性指数都表现为相同的规律:天然次生林显著高于两种人工林 (P<0.05),两种人工林之间差异不显著(P>0.05,表2)。
灌木层的物种丰富度在森林恢复方式之间的差异表现为天然次生林高于湿地松人工林,高于马尾松人工林,其中天然次生林与马尾松人工林的差异达到显著水平(P< 0.05,表2)。
3种森林恢复方式之间的草本层物种数没有显著差异,但其多样性指数表现为天然次生林显著高于两种人工林,两种人工林之间差异不显著(P>0.05,表2)。
2.4 植物群落结构相似性分析
调查到的所有323种植物物种用于植物群落结构相似性分析。DCA可以在二维图上较好的反映不同样地植物群落结构的差异。本文中,DCA能较好的区分3种森林恢复方式的植物群落结构(图2)。第一轴解释植物群落结构7.4%的变异,第二轴解释6.5%的变异。第一轴上的得分能显著地区分天然次生林与高于两种人工林,在第一轴上的得分表现为天然次生林显著高于两种人工林(P<0.05)。第二轴能显著区分湿地松人工林与其它两种森林恢复方式,在第二轴上的得分表现为湿地松人工林显著低于其它两种森林恢复方式(P<0.05)。在DCA图中,相比湿地松人工林天然次生林样地与马尾松人工林样地距离更近。这意味着相比湿地松人工林,天然次生林与马尾松人工林的植物群落结构相似性更大。

图1 3种森林恢复方式的植物区系构成Fig.1 Flora composition under 3 forest restoration approaches

表2 不同森林恢复方式植物群落多样性Table 2 Diversity of plant community under different forest restoration approaches
与第一轴显著正相关的物种有千年桐(Vernicia montana Lour)和紫金牛(Ardisia japonica);与第一轴显著负相关的物种有湿地松、紫珠(Callicarpa bodinieri)和构树(Broussonetia papyifera)。与第二轴显著负相关的物种为铁芒萁(Dicranopteris linearis);与第二轴显著正相关的物种为粗糠柴(Mallotus philippensis)、水青冈(Fagus longipetiolata)、子凌蒲桃(Syzygium championii)、米念芭(Tirpitzia ovoidea)、烟斗柯(Lithocarpus corneus)、三脉叶荚蒾(Viburnum triplinerve)、朴树(Celtis sinensis)、求米草(Oplismenus undulatifolius)和江南卷柏(Selaginella moellendorffii)。以上所有树种与DCA轴的相关系数均大于0.5。DCA图表明在天然次生林中千年桐和紫金牛分布较多;在湿地松人工林中湿地松、紫珠和构树分布较多。
多元方差分析表明,3种森林恢复方式的植物群落结构之间存在显著差异(表3,P=0.001)。恢复方式能解释湿地松人工林和天然次生林之间植物群落结构19.7%的变异,能解释马尾松人工林和天然次生林植物群落结构之间32.6%的变异,能解释湿地松人工林和马尾松人工林植物群落之间6.15%的变异。根据基于距离矩阵的方差分析(Adonis)中森林恢复方式解释量的大小以及相似性分析(ANOSIM)中的R值可知,相比湿地松人工林,马尾松人工林与天然次生林的植物群落结构更相似。
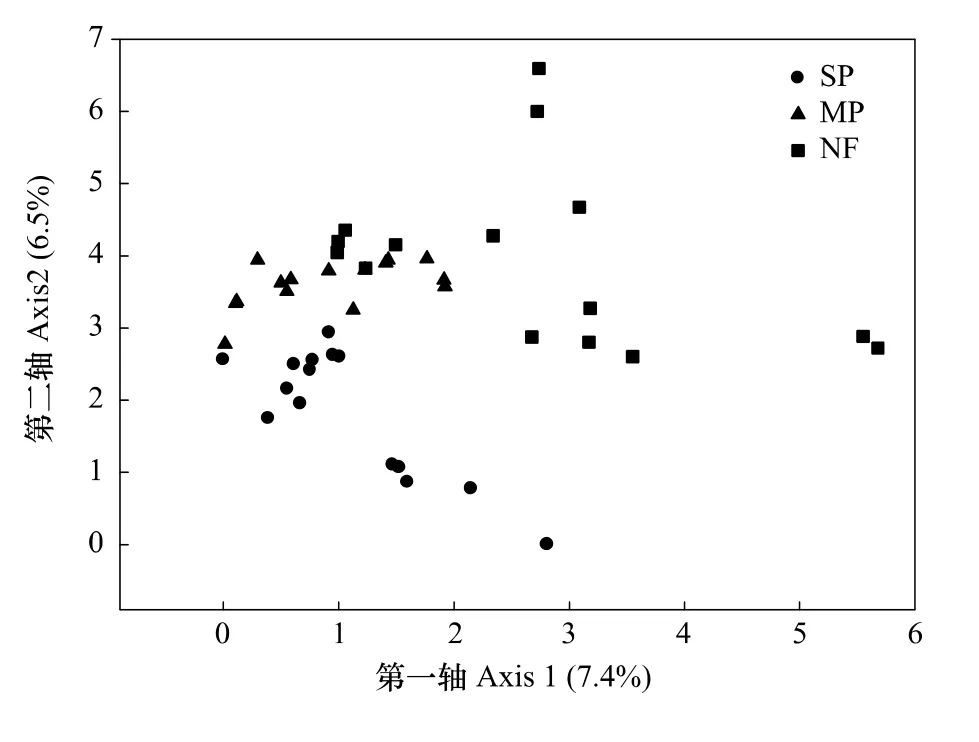
图2 3种森林恢复方式的植物群落DCA二维排序图Fig.2 DCA biplotofplantcommunityunder 3forest restoration approaches
3 讨论
本研究在天然次生林中调查到86科160属226种植物,高于在南方红壤区深圳的天然次生林中的植物21—64种[16],这与取样面积大小及空间分布有关,也表明本研究的植物群落调查更有代表性。
3.1 森林恢复方式对植物群落多样性的影响
植物物种多样性是森林恢复过程中群落变化的重要指标[10],多样性越高,生态服务功能越高[7,25-26],抗干扰能力越强[26-27]。天然次生林的物种丰富度、辛普森指数和植物区系基本构成和地理区系类型高于马尾松人工林和湿地松人工林,显示其物种的生态位宽度更宽、资源利用程度更高。前人的研究也有类似的报道,人工林的生物多样性远低于天然混交林[9],人工造林对提升生物多样性的贡献较小[6]。而小流域治理20 a后千烟洲的植物群落表现为天然次生林的乔木丰富度高于人工林,灌木层和草本层的植物多样性低于天然次生林,可能与其研究区的天然次生林受到的干扰较大,或者天然次生林的较高的乔木层郁闭度导致较低的林下植被多样性有关[28]。这与退化群落的属数以及温带成分比例低于干扰较小的常绿阔叶林[29]的趋势类似。天然次生林有更多的区系类型及变型,其优势植物主要来自壳斗科、山茶科和樟科,而这些物种可能发展成为亚热带植物群落的顶极植物种,表明天然次生林处于更高级的群落演替阶段[30]。

表3 植物群落结构的多元方差分析Table 3 Multi-variant analysis of variance based on plant community
本地种马尾松人工林的植物物种丰富度辛普森指数略高于引进种湿地松人工林。前人也曾报道,湿地松减少林下本地种的多样性[31]。这可能是由环境异质性和人为干扰的共同作用形成的结果。
3.2 不同森林恢复方式的植物群落结构相似性
DCA和多元方差分析都表明,恢复方式显著改变了植物群落结构。与天然次生林相比,人工林植物群落结构已发生显著改变,这表明人工造林改变了原初的植物群落结构,这会造成与原有生态系统的群落结构不匹配[8]。DCA图中湿地松人工林样方与天然次生林样方距离小于马尾松人工林样方与天然次生林样方距离,多元方差分析以及优势植物区系构成的结果都表明,湿地松人工林比马尾松人工林有更高的改变物种组成的能力。另外,湿地松人工林湿地松的重要值远远大于马尾松人工林中马尾松的重要值,说明相比马尾松,湿地松使群落单一化的能力更强。因而,使用外来种形成的先锋林可能会阻碍生态系统向稳定的地带性植被发展[4,32],本地种种植在景观管理[31]和森林恢复[33-34]中被推崇,以使生态风险最小化。
3.3 对森林恢复的启示
本文在我国南方红壤区综合乔木、灌木和草本的群落结构和多样性认为,自然恢复的天然次生林的植物多样性高于人工林,而本地种马尾松人工林又优高于引进种湿地松林人工林。因此在我国南方红壤区的植物群落多样性维持功能上,自然恢复优于人工恢复,而人工恢复中又以本地种马尾松人工林优于引进种湿地松。因而,在生态恢复过程中,应该提倡自然恢复[9,15],减少人为干扰[10]。Chazdon[8]则认为森林生态系统的恢复方式的选择取决于土壤和森林恢复的程度,残留的植被以及恢复目标,即应综合考虑物理条件、生物多样性的临界水平和可提供种源的景观基底值[4,35]和林下植被发展能力[33]。
综合以上研究认为,在我国南方红壤区,恢复方式对植物群落多样性有显著影响。自然恢复的天然次生林植物群落多样性高于人工恢复的马尾松人工林和湿地松人工林,本地种马尾松人工林在维持区域群落结构功能上优于引进种湿地松人工林。
致谢:湖南省林科院李锡泉研究员、中国科学院广西植物研究所刘演研究员和衡阳县林业局蒋大力在植物群落调查中给予指导,赵娟娟对写作给予帮助,黄志刚、陈红兴、王顺亮、马祝才、蒋日红、曾祥铭、吾望辉、刘胜福、农新东、黄愈松、蒋德龙、陈菲、李佳在植物群落调查中给予帮助,特此致谢。
[1]Ouyang Z Y,Wang X K,Miao H.A primary study on Chinese terrestrial ecosystem services and their ecological-economic values.Acta Ecologica Sinica,1999,19(5):607-613.
[2]Costanza R,d'Arge R,de Groot R D,Farber S,Grass M,Hannon B,Limburg K,Naeem S,O'Neil R V,Paruelo J,Raskin R G,Sutton P,Van den Belt M.The value of the world's ecosystem services and natural capital.Nature,1997,387(6630):253-260.
[3]Zhao T Q,Ouyang Z Y,Zheng H,Wang X K,Miao H.Forest ecosystem services and their valuation in China.Journal of Natural Resources,2004,19(4):480-491.
[4]Ren H,Li Z A,Shen W J,Yu Z Y,Peng S L,Liao C H,Ding M M,Wu J G.The changes of biodiversity and ecosystem services in forest restoration process in Southren China.Science in China Series C:Life Sciences,2006,36(6):563-569.
[5]Liu G H,Fu B J,Chen L D,Guo X D.Characteristics and distributions of degraded ecological types in China.Acta Ecologica Sinica,2000,20(1):13-19.
[6]Zheng H,Ouyang Z Y,Wang X K,Fang Z G,Zhao T Q,Miao H.Effects of regenerating forest cover on soil microbial communities:a case study in hilly red soil region,Southern China.Forest Ecology and Management,2005,217(2/3):244-254.
[7]Benayas J M R,Newton A C,Diaz A,Bullock J M.Enhancement of biodiversity and ecosystem services by ecological restoration:a meta-analysis.Science,2009,325(5944):1121-1124.
[8]Chazdon R L.Beyond deforestation:restoring forests and ecosystem services on degraded lands.Science,2008,320(5882):1458-1460.
[9]Sayer J,Chokkalingam U,Poulsen J.The restoration of forest biodiversity and ecological values.Forest Ecology and Management,2004,201(1):3-11.
[10]Zhang J T,Dong Y R.Factors affecting species diversity of plant communities and the restoration process in the loess area of China.Ecological Engineering,2010,36(3):345-350.
[11]Mo J M,Peng S L,Brown S,Kong G H,Fang Y T.Response of biomass production to human impacts in a pine forest in subtropical China.Acta Ecologica Sinica,2004,24(2):193-200.
[12]Xie J S,Guo J F,Yang Z J,Huang Z Q,Chen G S,Yang Y S.Rapid accumulation of carbon on severely eroded red soils through afforestation in subtropical China.Forest Ecology and Management,2012,doi:10.1016/j.foreco.2012.06.038.
[13]Tian D L,Xiang W H,Yan W D.Comparison of biomass dynamic and nutrient cycling between Pinus massomiana plantation and Pinus elliottii plantation.Acta Ecologica Sinica,2004,24(10):2207-2210.
[14]Zhang T P,Ren H,Peng S L,Yu Z Y.The ecological and biological characteristics of Pinus elliotti.Ecologic Science,1999,18(2):8-12.
[15]Zheng H,Chen F L,Ouyang Z Y,Tu N M,Xu W H,Wang X K,Miao H,Li X Q,Tian Y X.Impacts of reforestation approaches on runoff control in the hilly red soil region of Southern China.Journal of Hydrology,2008,356(1/2):174-184.
[16]Wang D P,Ji S Y,Chen F P,Xing F W,Peng S L.Diversity and relationship with succession of naturally regenerated southern subtropical forests in Shenzhen,China and its comparison with the zonal climax of Hong Kong.Forest Ecology and Management,2006,222(1/3):384-390.
[17]Bu Y J,Wen Z M,Jiao F,Jiao J Y.Research on bio-diversity of artificial and natural plant communities in loess hilly region-taking Ansai County as an example.Research of Soil and Water Conservation,2005,12(1):4-6.
[18]Wang Y,Ouyang Z Y,Zheng H,Wang X K,Chen F L,Zeng J.Carbon metabolism of soil microbial communities of restored forests in Southern China.Journal of Soils and Sediments,2011,11(5):789-799.
[19]Li Q H,Yang L W,Zhou Q X.Comparative analysis on species diversity of hillclosed afforested plant community in Beijing Jiulong Mountain.Chinese Journal of Applied Ecology,2002,13(9):1065-1068.
[20]Fang J Y,Wang X P,Shen Z H,Tang Z Y,He J S,Yu D,Jiang Y,Wang Z H,Zheng C Y,Zhu J L,Guo Z D.Methods and protocols for plant community inventory.Biodiversity Science,2009,17(6):533-548.
[21]Wu Z Y.The areal-types of Chinese genera of seed plants.Acta Botanica Yunnanica,1991,4(Suppl IV):1-139.
[22]Simpson E H.Measurement of diversity.Nature,1949,163:688.
[23]Zhao J J,Ouyang Z Y,Zheng H,Zhou W Q,Wang X K,Xu W H,Ni Y M.Plant species composition in green spaces within the built-up areas of Beijing,China.Plant Ecology,2009,209(2):1-16.
[24]Oksanen J,Kindt R,Legendre P,O'Hara B,Simpson G L,Solymos P,Henry M,Stevens H,Wagner H,2008.Vegan:community ecology package.R package version 1.15-1.http://cran.r-project.org/.http://vegan.r-forge.r-project.org/.http://cc.oulu.fi/wjarioksa/opetus/metodi/vegantutor.pdf(accessed 23.03.10)
[25]Malcolm D C,Mason W L,Clarke G C.The transformation of conifer forests in Britain-regeneration,gap size and silvicultural systems.Forest Ecology and Management,2001,151(1/3):7-23.
[26]Worm B,Barbier E,Beaumont N,Duffy J,Folke C,Halpern B S,Jackson J B C,Lotze H K,Micheli F,Palumbi S R,Sala E,Selkoe K A,Stachowicz J J,Watson R.Impacts of biodiversity loss on ocean ecosystem services.Science,2006,314(5800):787-790.
[27]Marcot B G.Biodiversity and the lexicon zoo.Forest Ecology and Management,2007,246(1):4-13.
[28]Liu J Q,Hu L L,Li X R.Plant diversity in Qianyanzhou after 20 years of small watershed treatment.Acta Phytoecologica Sinica,2005,29(5):766-774.
[29]Wang X H,Yan E R,Yan X,Wang L Y.Analysis of degraded evergreen broad-leaved forest communities in Eastern China and issues in forest restoration.Acta Ecologica Sinica,2005,25(7):1796-1803.
[30]Peng S L.Forest Community Dynamics in South-subtrophical Area.Beijing:Scientific Press,1996.
[31]Hedman C W,Grace S L,King S E.Vegetation composition and structure of southern coastal plain pine forests:an ecological comparison.Forest Ecology and Management,2000,134(1/3):233-247.
[32]Holl K D.Long-term vegetation recovery on reclaimed coal surface mines in the eastern USA.Journal of Applied Ecology,2002,39(6):960-970.
[33]Parrotta J A,Turnbull J W,Jone N.Catalyzing native forest regeneration on degraded tropical lands.Forest Ecology and Management,1997,99(1/2):1-7.
[34]Ma J M,Liu S R,Shi Z M,Liu X L,Miao N.A review on restoration evaluation studies of degraded forest ecosystem.Acta Ecologica Sinica,2010,30(12):3297-3303.
[35]Yang Y S,Guo J F,Wang J,Sheng H,Li X B,Lin C F,Wang W Q,Xie J S,Chen G S.Review on plant ecology in Southern China.Acta Geographica Sinica,2009,64(9):1048-1057.
参考文献:
[1]欧阳志云,王效科,苗鸿.中国陆地生态系统服务功能及其生态经济价值的初步研究.生态学报,1999,19(5):607-613.
[3]赵同谦,欧阳志云,郑华,王效科,苗鸿.中国森林生态系统服务功能及其价值评价.自然资源学报,2004,19(4):480-491.
[4]任海,李志安,申卫军,余作岳,彭少麟,廖崇惠,丁明懋,邬建国.中国南方热带森林恢复过程中生物多样性与生态系统功能的变化.中国科学C辑:生命科学,2006,36(6):563-569.
[5]刘国华,傅伯杰,陈利顶,郭旭东.中国生态退化的主要类型、特征及分布.生态学报,2000,20(1):13-19.
[11]莫江明,彭少麟,Brown S,孔国辉,方运霆.鼎湖山马尾松林群落生物量生产对人为干扰的响应.生态学报,2004,24(2):193-200.
[13]田大伦,项文化,闫文德.马尾松与湿地松人工林生物量动态及养分循环特征.生态学报,2004,24(10):2207-2210.
[14]张太平,任海,彭少麟,余作岳.湿地松(Pinus elliottii Engelm.)的生态生物学特征.生态科学,1999,18(2):8-12.
[17]卜耀军,温仲明,焦峰,焦菊英.黄土丘陵区人工与自然植物群落物种多样性研究——以安塞县为例.水土保持研究,2005,12(1):4-6.
[19]李清河,杨立文,周金星.北京九龙山植物群落物种多样性特征对比分析.应用生态学报,2002,13(9):1065-1068.
[20]方精云,王襄平,沈泽昊,唐志尧,贺金生,于丹,江源,王志恒,郑成洋,朱江玲,郭兆迪.植物群落清查的主要内容、方法和技术规范.生物多样性,2009,17(6):533-548.
[21]吴征镒.中国种子植物属的分布区类型.云南植物研究,1991,4(增刊IV):1-139.
[28]刘琪璟,胡理乐,李轩然.小流域治理20年后的千烟洲植物多样性.植物生态学报,2005,29(5):766-774.
[29]王希华,闫恩荣,严晓,王良衍.中国东部常绿阔叶林退化群落分析及恢复重建研究的一些问题.生态学报,2005,25(7):1796-1803.
[30]彭少麟.南亚热带森林群落动态学.北京:科学出版社,1996.
[34]马姜明,刘世荣,史作民,刘兴良,缪宁.退化森林生态系统恢复评价研究综述.生态学报,2010,30(12):3297-3303.
[35]杨玉盛,郭剑芬,王健,盛浩,李熙波,林成芳,王维奇,谢锦升,陈光水.我国南方植物生态学研究述评.地理学报,2009,64(9):1048-1057.
