植物叶片水稳定同位素研究进展
2013-08-02余武生万诗敏
罗 伦,余武生 ,万诗敏,周 平
(1.中国科学院青藏高原环境变化与地表过程重点实验室,中国科学院青藏高原研究所,北京 100101;2.中国科学院研究生院,北京100049;3.天津城市建设学院,基础学科部,天津 300384;4.长春气象探测中心,长春 130062)
植物叶片不仅是联系分子生态学和宏观生态学的重要桥梁,同时也是植物光合作用和呼吸作用进行物质和能量交换的重要场所。因此,植物叶片水稳定同位素变化可以直接沟通植物叶片内部与外界的物质和能量联系[1-2]。尤其是,通过研究植物叶片水稳定同位素可以反映植物生长周围的气象与生态信息,如温度[3]、相对湿度[4]、气压[5]和蒸腾速率[6]以及气孔导度等[7]。早在 1965 年,Gonfiantini等[8]就发现一些植物叶片水相对于植物水源会发生δ18O富集。随后,植物叶片水δD的富集现象也被发现[9]。进一步研究证实,不同物种植物叶片水相对于植物水源(土壤水或茎水)均会发生稳定同位素分馏[10-11]。特别是,一些学者将研究开放水体蒸发过程时的Craig-Gordon模型进行修正并应用到植物叶片水上[12-14],使得植物叶片水稳定同位素研究得到进一步拓展。而在模型的发展与应用过程中,又促进了对植物叶片水稳定同位素分馏机制及其与生态因子的相互关系的了解[1,15]。
植物叶片水作为参与水循环的一个重要环节,了解叶片水稳定同位素与大气水汽、降水、土壤水等不同水体中稳定同位素的相互关系,有助于揭示其在局地水体稳定同位素循环中的分配与贡献。而在实际研究中,大气水汽[16]、降水[17]、地表径流[18]、湖水与土壤水[19]等不同圈层的多相态水体稳定同位素研究相对成熟,对于植物圈的叶片水稳定同位素与水循环相结合的研究却相对较少,直到最近才逐渐受到科学家们的青睐。如,Lai等[20]曾在美国华盛顿州的南部森林建立了包含大气水汽、降水、土壤水、植物叶片水、茎水以及在稳态和非稳态环境下经过蒸腾而进入空气中的水汽等不同水体中稳定同位素水文循环。
在国内,稳定同位素生态学研究起步较晚[21-22],且大多数是关于确定植物水源[23-24]、水分利用[25]和生态效应[26]以及区分生态系统蒸散组分[27]等方面的研究,较少涉及到植物叶片水稳定同位素研究。近年来,该方面的研究逐渐得到重视:温学发等[28]介绍了叶片水δ18O的富集和大气水汽δ18O的原位观测技术;Zhang等[29]用稳定同位素技术区分生态系统中的蒸散发;特别是张世春揭示了冬小麦叶片水稳定同位素的日变化过程,发现叶片水稳定氢、氧同位素均表现为在6:00最低,12:00—15:00区间最高的变化趋势[30]。
随着国内植物叶片水稳定同位素研究逐渐展开,相关成果将逐渐得以报道。根据目前国内外研究进展,本文详细介绍了植物叶片水稳定氢、氧同位素在植物体中的分馏过程及形式,分析了叶片水稳定同位素组成与气象和生态因子的关系,阐述了叶片水稳定同位素相关模型,并讨论了这些模型可能存在的问题以及未来植物叶片水稳定同位素的研究趋势。
1 植物叶片水稳定同位素分馏过程及形式
植物叶片水稳定同位素组成一般用δL或ΔL表示:

式中,RL为叶片水中稳定同位素比值(18O/16O或D/H);RVSMOW为“标准平均大洋水”稳定同位素比值。

式中,ΔL表示叶片水相对于植物水源的稳定同位素组成;Rs为植物水源(茎水或土壤水)稳定同位素比值。
植物叶片水分来源于植物根系吸收土壤中的水,然后经过茎中导管传输到叶柄,最后进入植物叶片中。除泌盐类植物外[31],一般情况下,水分在到达叶片之前均不会发生稳定同位素分馏。当水分到达叶片时,由于分馏效应,植物叶片水稳定同位素在叶片上的分布并不均匀。研究发现,从叶片基部(叶基)沿叶脉向叶尖δD值明显升高,而且叶肉中的δD比叶脉、叶柄以及植物茎中的δD值均高[32]。δ18O同样也存在类似的趋势,由叶基向叶尖,叶脉向叶片边缘δ18O值逐渐升高[33]。在自然条件下,植物叶片水稳定同位素会发生热力学平衡分馏、动力学分馏和植物生化分馏3种形式的稳定同位素分馏。正是由于上述诸种分馏效应的存在导致叶片水稳定同位素在叶片各部位分布不均。
当水分到达叶片时,叶片水会发生蒸发,此时植物叶片水由液态变为气态,在此相变过程中会发生热力学平衡分馏效应。根据Majoube[34]的研究,与温度相关的平衡分馏系数可表示为:
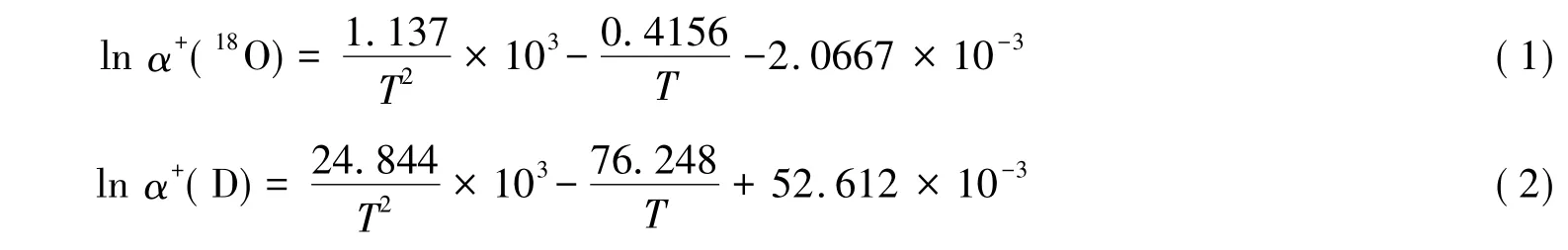
式中,α+表示平衡分馏系数,其与平衡分馏效应ε+的关系为:ε+=1000(α+-1)≈1000 lnα+(‰)(α+趋近于1时);T表示叶片的开尔文绝对温度。
植物叶片水除了会发生热力学平衡分馏外,水分子在穿过气孔进入大气过程中,需要克服一定的阻力,而轻、重同位素的扩散速率又不同,因此会发生动力学分馏。在此过程中,动力分馏效应(εk)与水分子扩散过程中受到的叶片气孔阻力(rs)和边界层阻力(rb)之间存在如下关系[35-37]:

式中,αks表示水分子穿过气孔时的动力分馏系数,αkb表示水分子穿过叶片边界层时的动力分馏系数,它们之间的关系为 αkb=αks2/3[10]。Merlivat[36]计算出 αks是 1.025(H/D) 和 1.0285(16O/18O),此时相应的 αkb为1.017(H/D)和 1.0189(16O/18O)[13-14]。2003 年 Cappa 等[37]将 αks进一步修正为 1.016(H/D)和 1.032(16O/18O)。
另外,植物进行生理活动时,也会发生分馏。比如,植物进行光合作用时,空气中的CO2作为重要反应物,通过气孔进入叶片的CO2只有部分被固定:在与水合成葡萄糖的过程,羰基氧与H2O分子中氧原子发生交换,使得δ18O发生分馏[38];而未被吸收的部分CO2与叶片水在碳酸酐酶(CA)催化下,经过稳定同位素平衡反应后返回大气,此时是以C18O16O的形式存在于大气中[39-40],同样发生δ18O分馏。而光合作用过程中摄取的氢参与合成有机物会发生一系列生物化学反应,这些生化反应会引起δD发生明显的负同位素分馏效应[41]。另外,在一些脂类物质的合成中,也会发生生化分馏[3]。虽然植物叶片水存在生化分馏效应,但是与热力学平衡分馏效应和动力学分馏效应相比,其分馏过程复杂。故本文未重点涉及生化分馏效应。
2 植物叶片水稳定同位素与气象、生态因子的关系
2.1 气象因子
在植物叶片水稳定同位素的平衡分馏过程中,其分馏程度受制于温度的影响:随着温度升高,平衡分馏效应降低。目前关于叶片水δ18O和δD与温度关系的研究结论不尽相同。Dongmann等[10]研究山毛榉(Fagus silvatica)和橡木(Quercus robur)叶片水δ18O发现,其与温度之间存在很好的正相关性。Zhou等[3]选取3种C3植物和3种C4植物分别在20℃和30℃时进行对比研究,结果显示,在30℃时所有植物叶片水δD的值均比在20℃时的值大,即高温有助于叶片水δD的富集。但是,Cooper和DeNiro[11]在加州大学洛杉矶分校校园(UCLA)和棕榈沙漠镇的峡谷研究中心(Deep Canyon)两个地区研究5种C3植物和4种CAM植物叶片水δ18O和δD与温度的关系时,发现仅在Deep Canyon干旱地区仅两种C3植物叶片水的δ18O与温度呈正相关性,Deep Canyon干旱地区的其它植物和UCLA地区所有植物的叶片水δ18O和δD与温度之间均无统计意义。这说明,不同地区不同植物种类的叶片水稳定氢、氧同位素对温度因子的响应并不一致。
相对湿度(h)是影响植物叶片水稳定同位素富集的一个重要因子。相对湿度对叶片水稳定同位素组成的影响是由大气水汽压(ea)和叶内细胞间水汽压(ei)共同决定的,其关系为h=ea/ei[1]。当湿度增加,叶内外水汽压差变小,植物蒸腾降低,叶片水富集作用减小[42],即相对湿度与叶片水稳定同位素之间存在负相关性。比如,干季比湿季植物叶片水更富集[4,43];白天比夜间植物叶片水更富集[44]。此外,研究发现相对湿度越高,相对湿度与叶片水稳定同位素之间的相关性更为显著[45-46]。这可能与叶片自身空间异质性有关:相对湿度越大,异质性越小;相对湿度越小,异质性越大[46]。有研究证实,相对湿度越低,叶尖与叶基两处的Δ18O差异越大[47-48]。同样地,Šantr˚ucˇek等[32]发现ΔD也表现出类似的现象。
气压的变化导致植物蒸腾速率的改变,因而影响动力分馏过程,进而也影响叶片水的富集。不过,很少直接探讨大气气压对植物叶片水稳定同位素富集的影响,更多的是利用叶-气水汽压差(leaf-to-air vapor pressure difference,叶内水汽压与大气水汽压的差值)或空气水汽压亏缺(atmospheric vapor pressure deficit,空气饱和水汽压与实际水汽压的差值)研究其对叶片水稳定同位素富集的影响。如Ripullone等[7]对棉花叶片水研究发现,在气温为29℃时,叶片水Δ18O与叶-气水汽压差存在正相关;当温度为20℃时,叶片水Δ18O与叶-气水汽压差的相关性增强。另外,Li等[5]通过对落叶松的研究证实了叶片水Δ18O与空气水汽压亏缺之间存在显著的正相关。
风对蒸腾作用的影响取决于风速。微风能把气孔外面的水蒸气吹走,促使植物进一步蒸腾,但是强风可能引起气孔关闭,植物蒸腾变慢[49]。因而,风速对植物叶片水稳定同位素富集的影响较为复杂,导致二者之间的相关性难以显著[10]。不过,在森林水汽研究中,冠层上端的水汽δ18O与风速之间存在一定的联系[20]。由于水汽δ18O与植物水源δ18O存在分馏[50],而水源δ18O又是叶片水δ18O的初始来源,故风速通过影响水汽δ18O而间接影响叶片水稳定同位素组成。
2.2 生态因子
植物在蒸腾过程中,水汽会逃逸出叶片,从而导致叶片水相对富集,相对而言,蒸腾出去的水汽δT相对贫化。蒸腾作用的大小受外界环境和内部阻力共同影响。植物叶片水稳定同位素组成与蒸腾速率(E)之间存在显著的正相关。如,Sheshshayee等[6]在研究向日葵叶片水Δ18O时就发现这种相关性。另外,叶片各部位蒸腾速率的不均一性,导致叶片各部位富集程度不同。如,Šantr˚ucˇek等[32]发现雪花桉(Eucalyptus pauciflora)叶片无论在干旱或湿润环境下,叶基的蒸腾速率小于叶尖的,因而叶基的ΔD值小于叶尖的。
气孔导度gs通过影响植物叶片的蒸腾,进而对动力分馏过程产生一定的影响。如果其它因子的影响不变,气孔导度gs变大时,则气孔阻力rs减小,植物蒸腾速率增大,动力分馏效应相应变化。Sheshshayee等[6]指出转基因豇豆叶片水Δ18O与gs之间呈正相关。而Ripullone等[7]发现,在温度为20℃时,棉花叶片水Δ18O与gs二者之间存在显著的负相关,而当温度为29℃时,这种相关性稍微变弱。这种关于气孔导度对叶片水稳定同位素的不同影响的报道,可能与植物种类的不同有关。
此外,叶片长度[51]、水分胁迫[52]、露水[53]等其它生态因子以及光合作用[39-41]等植物生理过程都会影响植物叶片水稳定同位素组成。
3 植物叶片水稳定氢、氧同位素关系及意义
早在1961年,Craig[54]发现全球大气降水中δ18O和δD有很好的线性关系:δD=8δ18O+10。植物果汁中δ18O和δD也存在很好的线性关系,但是其斜率却比当地大气降水线的斜率低。比如,Bong等[55]研究西红柿的果汁中δ18O和δD,发现其线性方程的斜率仅为2.52。Allison等[56]发现植物叶片上每一个位置的δ18O和δD之间也存在一定的线性关系,并将其称为叶片水蒸腾线,定义为:
δD=mδ18O+b (4)式中,m的数学意义表示回归线的斜率,b表示回归线的截距。
叶片水蒸腾线的斜率m显著低于当地降水线的斜率,但稍微高于蒸发点水汽δD与δ18O回归线的斜率。比如Cernusak等[57]拟合了狭叶羽扇豆(Lupinus angustifolius)叶片水 δD与 δ18O之间的回归方程为δD=3.1δ18O-6.4‰,而当地降水线方程为 δD=7.1δ18O+11.1‰;Kim 和 Lee[2]的研究表明棉花叶片水蒸腾线的斜率为3.0,蒸发点水汽δD与δ18O回归线的斜率为2.6。目前研究结果中,叶片水蒸腾线的斜率m均较小。Cooper和DeNiro[11]研究发现9种植物叶片水蒸腾线的斜率从1.5到3.21不等,斜率的波动受当地气温和湿度的共同影响,而且斜率较小的植物叶片水稳定同位素与当地的气温和相对湿度的相关性更显著。Yakir等[58]也发现叶片水蒸腾线的斜率m可能受相对湿度影响:在较湿润(平均相对湿度59%)条件下,棉花叶片水蒸腾线的斜率为2.4;在较干燥条件(平均相对湿度45%)下,棉花叶片水蒸腾线的斜率为2.0。如果在干旱或半干旱条件下,斜率m会更低。如Gat等[59]发现在西亚干旱区正午时,多种植物叶片水蒸腾线的平均斜率在2.1到2.8之间,而在半干旱条件下,松针叶片水蒸腾线的斜率仅为1.3[56]。
在相同条件下,H182O相对于H162O发生的动力分馏效应大于DH16O相对于H162O的[17,60],故叶片水蒸腾线中斜率m越低,说明在分馏过程中动力分馏比平衡分馏的作用更大[60]。因此叶片水蒸腾线的斜率的大小可以指示叶片水稳定同位素动力分馏的强弱程度[2]。
4 叶片水稳定同位素主要模型
4.1 修正的Craig-Gordon稳态模型
由于直接测定植物叶片水稳定同位素相对复杂且费用较高,不能满足大量实验的数据要求。因此,通过一些比较容易测定的参数建立模型来确定植物叶片水稳定同位素值显得很有必要。早期,生态学家将Craig-Gordon模型进行修正应用于植物蒸腾过程稳定同位素模拟。该模型基于假设植物蒸腾δT与植物茎水δs相等。但是通过比较模拟预测值与实际测定值发现,二者之间存在一定的差异。一般情况下,修正的Craig-Gordon稳态模型预测值比实际值高[13-14,32]。但是也有研究发现,修正的Craig-Gordon稳态模型预测叶片水稳定同位素的平均值比实际观测值的平均值偏低[33,51]。其原因可能为:(1)该模型不能反映叶片水稳定同位素组成的叶片尺度空间的变化;(2)Craig-Gordon方程的稳态假设只对某一特定时段(比如,中午)或比较长的时间尺度(数天到数周)才有效[61]。
4.2 String-of-lakes模型
为了更好地模拟叶片水稳定同位素组成的叶片尺度的空间变化,string-of-lakes模型得以应运而生。该模型将叶片视为一个湖泊,叶基相当于湖泊的底部,叶尖相当于湖面。在湖水稳定同位素研究中,表层湖水由于蒸发导致稳定同位素富集,底层湖水蒸发小,则不容易富集。因而,从表层到底层,稳定同位素富集程度逐渐降低,形成一个梯度。同理,叶基到叶尖也会由于富集程度不同,产生一个梯度[62]。根据string-of-lakes模型,可把叶片从基部到叶尖等分成n段,每一部分的叶片水δ值是不同的,各段之间是呈梯度变化,然后分别求出各段的叶片水 δ值,第 n段叶片水 δn值可表示如下[51,62]:

式中,δa表示大气水汽稳定同位素组成;δn表示液态水进入第n段的蒸发面(比如气孔)时的δ值;F+和E分别代表流入和蒸发流出某一段的通量;h表示叶片周围的相对湿度;ε=ε++(1-h)εk,ε+为液-气平衡分馏效应,εk为动力分馏效应。在该模型中,先求解出各段的δ值,然后再对整个叶片求平均[51]。
4.3 Péclet效应的稳态模型——Farquhar& Lloyd模型
为了提高模型模拟的精确性,Farquhar和Lloyd[63]引入Péclet效应对Craig-Gordon模型进行修正。Péclet效应是指水在叶片中从叶脉到叶片边缘蒸发点迁移过程中,由于轻水较重水容易蒸发,故在蒸发点重水富集,从而导致重同位素又会从叶片边缘蒸发点逆向扩散到叶脉[52]。其原理先通过Craig-Gordon模型计算出蒸发点水的稳定同位素组成Δes(因其是不可测定的),可以近似表示为[35,63]:

式中,Δes、Δv分别表示叶片蒸发点水和大气水汽相对于植物水源的稳定同位素组成。ea、ei分别表示大气水汽压和叶内细胞间水汽压。在确定了 Δes后,可通过如下Péclet效应方程确定整个叶片水稳定同位素组成[63]:

式中,ΔLs为Péclet效应模型模拟的叶片水相对于植物水源的稳定同位素组成;Δes可以通过方程(6)得到;y为 Péclet数,其表达式为[63]:

式中,E表示蒸腾速率(mol·m-2·s-1);L表示叶脉到叶片边缘蒸发点的有效长度(m);C表示水的摩尔浓度(5.56×104mol/m3);Dw表示是示踪物(H182O或HDO)在水中的扩散率。
Péclet效应在Craig-Gordon模型上的应用,提高了模型模拟的精度[44,64]。但在野外条件下模拟效果并不理想[65],这是由于叶片各个部位的蒸发强度不同,叶片水运动的有效距离很难精确,而Péclet效应也仅考虑了横向(叶脉到蒸发点)一个方向上的变化。于是,有学者提出通过增加Péclet数以进一步校正Craig-Gordon模型,即混合路径 Péclet效应模型,公式如下[47]:

式中,Δx、Δrv和Δm分别表示经过Péclet效应校正后的叶片主脉、细叶脉以及叶肉等各组分水的Δ值;φx、φrv和φm表示对应的组分水在叶片水中的比重(φx+φrv+φm=1);ΔLs表示混合路径模型的最后模拟值。一般情况下,由于细叶脉的水所占比例少,可以忽略中间项。而Cuntz等[61]认为,叶脉(主脉)一般在叶片的中央,而叶片的向上、下表面都有气孔,都会导致叶片水的蒸发富集,且富集程度不同,故在稳态条件下需对叶片上、下表面分别进行Péclet效应校正。ΔLs可以表示为:

式中,V为整个叶片的含水量(mol/m2),Vup和Vdown分别表示叶上、下表面含水量(mol/m2),yup和ydown分别表示叶片上、下表面的Péclet数。
4.4 非稳态效应的模型——Dongmann模型
上述模型的建立均是在稳态条件下建立的,因此可以称为稳态模型。然而研究发现植物叶片水稳定同位素只有在正午时分才能接近稳态。因此,要得到理想的模拟效果,模型建立的时间因素不能忽略。代表性的模型为1974年Dongmann等[10]提出的一种简单的非稳态模型,该模型可以定量研究叶片水稳定同位素的分馏与富集。Cernusak等[57]人进一步将其完善如下:

式中,ΔLn表示非稳态模型计算出的叶片水相对于水源的稳定同位素组成;W表示为叶片水浓度(mol/m2);wi表示细胞间水蒸气的摩尔分数(mol/mol);g表示气孔导度和边界层导度之和(mol·m-2·s-1);αk为动力分馏系数;t表示发生稳定同位素分馏的时间(s)。
4.5 Péclet效应的非稳态模型——Farquhar& Cernusak模型
但Dongmann模型没有考虑Péclet效应。为了进一步提高模拟的精度,2005年Farquhar和Cernusak开发了一种同时考虑Péclet效应和非稳态效应的模型,其表示如下[65]:
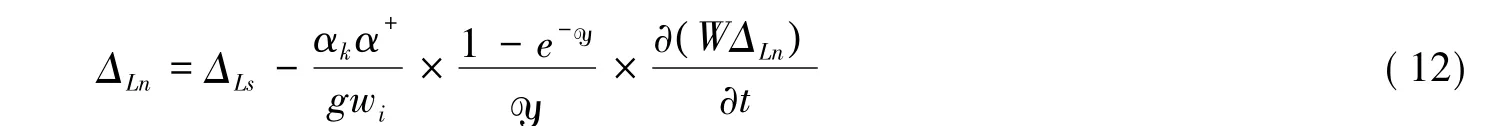
不过Cernusak等发现由于夜晚植物蒸腾速率小,y也就很小,因此在夜晚可以忽略Péclet效应[57]。故式(12)在夜间可以表示为方程(11)。
4.6 二维模型
近年来,一些学者开始尝试从二维的角度构建相关模型。2007年Ogée等[64]从叶片解剖学的空间模式角度探讨了稳定同位素在单子叶植物叶片的叶脉和叶肉中扩散的混合路径,提出了二维模型的概念。该模型很好地解释了C3和C4植物叶片水稳定同位素富集的差异是由于叶肉的曲折度引起,而不是简单地取决于叶脉与蒸发点之间的直线距离[64]。之后Shu等[66]对二维模型进行了部分修正并应用于美国北部的松针的研究,该模型重要的部分在于引进了蒸腾速率的累积函数(f),精确地模拟了松针从叶基到叶尖δ18O的富集情况。不过,由于该模型可能未充分考虑植物所有生物过程,所以对叶片水δD的模拟效果仍不理想。
5 问题与展望
尽管植物叶片水稳定同位素研究取得了长足的进展,但该领域的研究还存在很多问题,主要表现在以下几个方面:
在稳定同位素分馏与估算方面,目前叶片水稳定同位素平衡分馏的计算,主要是借鉴于其它水体稳定同位素平衡分馏机理。但是,植物叶片水稳定同位素分馏有其特殊性。比如,在植物叶片水中存在生化分馏,而其它水体几乎不存在这种分馏形式,而且目前尚不清楚植物叶片水中的生化分馏会导致多大程度的重同位素富集或亏损。另外,不同种类的植物叶片水稳定同位素分馏也较其它水体复杂。如,在计算植物叶片水动力分馏时,只考虑气孔阻力与边界层阻力,这意味着植物叶片的蒸腾,全部通过气孔进行。但是一些喜阴或喜湿的植物,角质层蒸腾是重要的部分,因此对于此种类的植物,需要考虑角质层阻力对稳定同位素分馏的影响。
在气象与生态因子方面,相对湿度、蒸腾速率、叶片内外水汽压差和水汽压亏缺等因子均与叶片水稳定同位素之间存在显著的正相关性。而温度与部分植物的叶片水稳定同位素之间存在一定的正相关,但在某些植物叶片水中二者的相关性并不明显[11]。另外,气孔导度与叶片水Δ18O的关系也不确定,这可能与植物种类有关,即不同的植物,气孔导度对叶片水Δ18O的影响可能不同。如Kodama等[67]研究发现单子叶植物黑小麦(Triticum×Secale)叶片水Δ18O主要受叶肉导度而不是气孔导度的影响。Ferrio等[15]认为葡萄(Vitis vinifera)叶片水Δ18O受气孔导度和水力导度共同作用。另外,一些环境因子(如风速、光照等)会改变蒸腾速率,但目前这些因子对叶片水稳定同位素的影响尚未发现统计学上的意义。
在稳定氢、氧同位素关系方面,很多报道已证实叶片水稳定氢、氧同位素之间存在线性相关,且叶片水蒸腾线的斜率指示叶片水稳定同位素的动力分馏[2]。而关于截距b的实际意义,鲜有报道。另外,在降水中过量氘d(d=δD-8δ18O)[17]是指示水汽来源的一项重要指标,而在植物叶片水稳定同位素研究中很少涉及过量氘及其意义的研究。
在模型方面,稳态模型中的修正的 Craig-Gordon模型[13-14]、string-of-lakes模型[62]、非稳态模型中的Dongmann模型[10]以及Cernusak等[57]人改进的非稳态模型均未考虑Péclet效应。稳态模型中的Farquhar&Lloyd 模型[63]、非稳态的Farquhar& Cernusak 模型[65]以及二维模型[66]均考虑了 Péclet效应。研究表明Péclet效应与 Craig-Gordon方程的耦合,虽然能够提高稳态模型模拟的精度[44,64],但其模拟的效果仍然不及Farquhar&Cernusak模型的理想[1]。string-of-lakes模型考虑了叶片水δ值从叶基到叶尖纵向的不同,故其对平行脉叶片的水稳定同位素模拟具有一定的优势,而对网状脉叶片的模拟效果较差[32-33,51]。二维模型能够提高δ18O的模拟精度[66],但该模型对δD的应用有待进一步完善。
因此,结合国内外的研究现状和存在的问题,需要从以下几个方面进行改进和拓展:
(1)加强植物叶片水稳定同位素在生理过程中变化的研究,明确不同生理过程中生化分馏效应对重同位素富集或亏损的影响。
(2)由于叶片水稳定同位素组成与温度、风速、气孔导度等气象、生态因子之间的关系仍然不够明确。因此,需要深入研究叶片水稳定同位素组成与这些要素之间的联系,进一步揭示叶片水稳定同位素富集的内在机制。
(3)加强叶片水中的δ18O和δD之间的线性关系研究,在已有的研究基础上,充分挖掘拟合曲线斜率m和截距b所代表的实际意义,并进一步揭示植物叶片水蒸腾线的斜率m远低于大气降水线的斜率的原因。另外,需要加强叶片水过量氘的实际意义的研究。
(4)当前模型的开发大多是在室内通过控制某些气象和生态因子模拟而成,因此需要通过野外实际测量,判定相关模型对不同气候区不同类型的植物的适用性,特别是在极端环境下(如,青藏高原等)的应用情况,并进一步完善相关模型。
(5)加强区域稳定同位素循环研究,揭示植物叶片水与当地大气降水、地表径流、土壤水、地下水以及大气水汽等不同水体稳定同位素的内在联系,从而构建区域稳定同位素的循环模式。
最后值得一提的是,随着技术的发展,近年来出现了稳定同位素激光分析仪(IRIS),这种新的同位素分析仪具有成本低、体积小、操作简单以及可实现原位测量等优点。但是稳定同位素激光分析仪在测量植物水或土壤水时,由于水样中有机污染物的存在会引起较大的误差[68]。为了减小这种误差,厂商发布了一种数据软件,而实际分析发现,该数据软件只能进行数据质量控制,无法消除有机物的干扰[69-70]。于是,最近又推出了一种微型高温热解模块技术,期望通过对有机物燃烧以消除有机物的存在对测量结果的影响。新技术的不断突破,对植物叶片水稳定同位素研究带来了测量的便捷和精度的提高,将推动植物叶片水稳定同位素研究的进一步发展。
[1]Kahmen A,Simonin K,Tu K,Goldsmith G R,Dawson T E.The influence of species and growing conditions on the 18-O enrichment of leaf water and its impact on‘effective path length’.New Phytologist,2009,184(3):619-630.
[2]Kim K,Lee X.Isotopic enrichment of liquid water during evaporation from water surfaces.Journal of Hydrology,2011,399(3/4):364-375.
[3]Zhou Y,Grice K,Chikaraishi Y,Stuart-Williams H,Farquhar G D,Ohkouchi N.Temperature effect on leaf water deuterium enrichment and isotopic fractionation during leaf lipid biosynthesis:Results from controlled growth of C3and C4land plants.Phytochemistry,2011,72(2/3):207-213.
[4]Cernusak L A,Mejia-Chang M,Winter K,Griffiths H.Oxygen isotope composition of CAM and C3Clusia species:non-steady-state dynamics control leaf water18O enrichment in succulent leaves.Plant,Cell and Environment,2008,31(11):1644-1662.
[5]Li S-G,Tsujimura M,Sugimoto A,Sasaki L,Yamanaka T,Davaa G,Oyunbaatar D,Sugita M.Seasonal variation in oxygen isotope composition of waters for a montane larch forest in Mongolia.Trees-Structure and Function,2006,20(1):122-130.
[6]Sheshshayee M S,Bindumadhava H,Ramesh R,Prasad T G,Lakshminarayana M R,Udayakumar M.Oxygen isotope enrichment(Δ18O)as a measure of time-averaged transpiration rate.Journal of Experimental Botany,2005,56(422):3033-3039.
[7]Ripullone F,Matsuo N,Stuart-Williams H,Wong S C,Borghetti M,Tani M,Farquhar G.Environmental effects on oxygen isotope enrichment of leaf water in cotton leaves.Plant Physiology,2008,146(2):729-736.
[8]Gonfiantini R,Gratziu S,Tongiorgi E.Oxygen isotopic composition of water in leaves//Isotopes and Radiation in Soil-Plant Nutrition Studies.Vienna:Proceedings of a Symposium,International Atomic Energy Agency,1965:405-410.
[9]Wershaw R L,Friedman I,Heller S J,Frank P A.Hydrogen isotopic fractionation of water passing through trees//Hobson G D,Speers G C,eds.Advances in Organic Geochemistry.Oxford:Pergamon Press,1970:55-67.
[10]Dongmann G,Nürnberg H W,Förstel H,Wagener K.On the enrichment of H182O in the leaves of transpiring plants.Radiation and Environmental Biophysics,1974,11(1):41-52.
[11]Cooper L W,DeNiro M J.Covariance of oxygen and hydrogen isotopic compositions in plant water:species effects.Ecology,1989,70(6):1619-1628.
[12]Craig H,Gordon L.Deuterium and oxygen-18 variations in the ocean and the marine atmosphere//Tongiorgi E,ed.Stable Isotopes in Oceanographic Studies and Paleotemperatures.Spoleto,1965:9-130.
[13]Flanagan L B,Ehleringer J R.Stable isotope composition of stem and leaf water:Applications to the study of plant water use.Functional Ecology,1991,5(2):270-277.
[14]Flanagan L B,Comstock J P,Ehleringer J R.Comparison of modeled and observed environmental influences on the stable oxygen and hydrogen isotope composition of leaf water in Phaseolus vulgaris L.Plant Physiology,1991,96(2):588-596.
[15]Ferrio J P,Pou A,Florez-Sarasa I,Cessler A,Kodama N,Flexas J,Ribas-Carbó M.The Péclet effect on leaf water enrichment correlates with leafhydraulic conductance and mesophyll conductance for CO2.Plant,Cell and Environment,2012,35(3):611-625.
[16]Wen X F,Zhang S C,Sun X M,Yu G R,Lee X.Water vapor and precipitation isotope ratios in Beijing,China.Journal of Geophysical Research,2010,115:D01103,doi:10.1029/2009JD012408.
[17]Dansgaard W.Stable isotopes in precipitation.Tellus,1964,16(4):436-468.
[18]Meredith K T,Hollins S E,Hughes C E,Cendón D I,Hankin S,Stone D J M.Temporal variation in stable isotopes(18O and2H)and major ion concentrations within the Daring River between Bourke and Wilcannia due to variable flows,saline groundwater influx and evaporation.Journal of Hydrology,2009,378(3/4):313-324.
[19]Gibson J J,Birks S J,Edwards T W D.Global prediction of δAand δ2H-δ18O evaporation slopes for lakes and soil water accounting for seasonality.Global Biogeochemical Cycles,2008,22:GB2031,doi:10.1029/2007GB002997.
[20]Lai C T,Ehleringer J R,Bond B J,Paw U K T.Contributions of evaporation,isotopic non-steady state transpiration and atmospheric mixing on the δ18O of water vapour in Pacific Northwest coniferous forests.Plant,Cell and Environment,2006,29(1):77-94.
[21]Wu S H,Pan T,Dai E F.The progress and prospect of stable isotopes in plants.Progress in Geography,2006,25(3):1-11.
[22]Lin G H.Stable isotope ecology:a new branch of ecology resulted from technology advances.Chinese Journal of Plant Ecology,2010,34(2):119-122.
[23]Duan D Y,Ouyang H,Song M H,Hu Q W.Water sources of dominant species in three alpine ecosystems on the Tibetan Plateau,China.Journal of Integrative Plant Biology,2008,50(3):257-264.
[24]Zhou C X,Sun Z Y,Yu S W.Using D and18O stable isotopes to determine the water sources of sand dune plants in Linze,middle reaches of Heihe River.Geological Science and Technology Information,2011,30(5):103-109.
[25]Sun S J,Meng P,Zhang J S,Huang H,Wan X C.Deuterium isotope variation and water use in an agroforestry system in the rocky mountainous area of North China.Acta Ecologica Sinica,2010,30(14):3717-3726.
[26]Sun S,Huang J,Han X,Lin G.Comparisons in water relations of plants between newly formed riparian and non-riparian habitats along the bank of Three Gorges Reservoir,China.Trees-Structure and Function,2008,22(5):717-728.
[27]Yuan G F,Zhang N,Sun X M,Wen X F,Zhang S C.Partitioning wheat field evapotranspiration using Keeling Plot method and continuous atmospheric vapor δ18O data.Chinese Journal of Plant Ecology,2010,34(2):170-178.
[28]Wen X F,Zhang S C,Sun X M,Yu G R.Recent advances in H182O enrichment in leaf water.Chinese Journal of Plant Ecology,2008,32(4):961-966.
[29]Zhang S C,Wen X F,Wang J L,Yu G R,Sun X M.The use of stable isotopes to partition evapotranspiration fluxes into evaporation and transpiration.Acta Ecologica Sinica,2010,30(4):201-209.
[30]Zhang S C.Continuous Measurements of Atmospheric Water Vapor δ18O and δD for Hydrological and Ecological Applications [D].Beijing:Graduate University of Chinese Academy of Sciences(Institute of Geographic Sciences and Natural Resources Research),2010.
[31]Lin G H,Sternberg S L.Hydrogen isotopic fractionation by plant roots during water uptake in coastal wetland plants//Ehleringer J R,Hall A E,Farquhar G D,eds.Stable Isotopes and Plant Carbon-Water Relations.San Diego:Academic Press,1993:497-510.
[32]Šantr˚ucˇek J,Kveˇtonˇek J,Šetlík J,Bulícˇková L.Spatial variation of deuterium enrichment in bulk water of snowgum leaves.Plant Physiology,2007,143(1):88-97.
[33]Gan K S,Wong S C,Yong J W H,Farquhar G D.18O spatial patterns of vein xylem water,leaf water,and dry matter in cotton leaves.Plant Physiology,2002,130(2):1008-1021.
[34]Majoube M.Fractionnement en oxygène 18 et en deutérium entre l'eau et sa vapeur.Journal de Chimie Physique et de Physico-Chimie Biologique,1971,10:1423-1436.
[35]Farquhar G D,Hubick K T,Condon A G,Richards R A.Carbon isotope fractionation and plant water-use efficiency//Rundel P W,Ehleringer J R,Nagy K A,eds.Stable Isotopes in Ecological Research.New York Springer,1988,68:21-40.
[36]Merlivat L.Molecular diffusivities of H216O,HD16O and H218O in gases.Journal of Physical Chemistry,1978,69(6):2864-2871.
[37]Cappa C D,Hendricks M B,DePaolo D J,Cohen R C.Isotopic fractionation of water during evaporation.Journal of Geophysical Research,2003,108(D16):4525-4534.
[38]Guy R D,Fogel M L,Berry J A.Photosynthetic fractionation of the stable isotopes of oxygen and carbon.Plant Physiology,1993,101(1):37-47.
[39]Gillon J S,Yakir D.Naturally low carbonic anhydrase activity in C4and C3plants limits discrimination against C18OO during photosynthesis.Plant,Cell and Environment,2000,23(9):903-915.
[40]Gillon J S,Yakir D.Influence of carbonic anhydrase activity in terrestrial vegetation on the18O content of atmospheric CO2.Science,2001,291(5513):2584-2587.
[41]Yakir D,DeNiro M J.Oxygen and hydrogen isotope fractionation during cellulose metabolism in Lemna gibba L.Plant Physiology,1990,93(1):325-332.
[42]Butt S,Ali M,Fazil M,Latif Z.Seasonal variations in the isotopic composition of leaf and stem water from an arid region of Southeast Asia.Hydrological Sciences Journal,2010,55(5):844-848.
[43]Lai C T,Ometto J P H B,Berry J A,Martinelli L A,Domingues T F,Ehleringer J R.Life form-specific variations in leaf water oxygen-18 enrichment in Amazonian vegetation.Oecologia,2008,157(2):197-210.
[44]Snyder K A,Monnar R,Poulson S R,Hartsough P,Biondi F.Diurnal variations of needle water isotopic ratios in two pine species.Trees-Structure and Function,2010,24(3):585-595.
[45]Kahmen A,Simonin K,Tu K P,Merchant A,Callister A,Siegwolf R,Dawson T E,Arndt S K.Effects of environmental parameters,leaf physiological properties and leaf water relations on leaf water δ18O enrichment in different Eucalyptus species.Plant,Cell and Environment,2008,31(6):738-751.
[46]Helliker B R.On the controls of leaf-water oxygen isotope ratios in the atmospheric crassulacean acid metabolism epiphyte Tillandsia usneoides.Plant Physiology,2011,155(4):2096-2107.
[47]Farquhar G D,Gan K S.On the progressive enrichment of the oxygen isotopic composition of water along a leaf.Plant,Cell and Environment,2003,26(6):801-819.
[48]Helliker B R,Ehleringer J R.Differential18O enrichment of leaf cellulose in C3versus C4grasses.Functional Plant Biology,2002,29(4):435-442.
[49]Li Q,Tan X L.Research advances in arid plant evapotranspiration.Gansu Agricultural Science and Technology,2006,12(10):18-20.
[50]Song X,Barbour M M,Saurer M,Helliker B R.Examining the large-scale convergence of photosynthesis-weighted tree leaf temperatures through stable oxygen isotope analysis of multiple data sets.New Phytologist,2011,192(4):912-924.
[51]Helliker B R,Ehleringer J R.Establishing a grassland signature in veins:18O in the leaf water of C3and C4grasses.Proceedings of the National Academy of Sciences of the United States of America,2000,97(14):7894-7898.
[52]Welp L R,Lee X,Kim K,Griffis T J,Billmark K A,Baker J M. δ18O of water vapour,evapotranspiration and the sites of leaf water evaporation in a soybean canopy.Plant,Cell and Environment,2008,31(9):1214-1228.
[53]Kim K,Lee X.Transition of stable isotope ratios of leaf water under simulated dew formation.Plant,Cell and Environment,2011,34(10):1790-1801.
[54]Craig H.Isotopic variations in meteoric water.Science,1961,133(3465):1702-1703.
[55]Bong Y S,Lee K S,Shin W J,Ryu J S.Comparison of the oxygen and hydrogen isotopes in the juices of fast-growing vegetables and slow-growing fruits.Rapid Communications in Mass Spectrometry,2008,22(18):2809-2812.
[56]Allison G B,Gat J R,Leaney F W.The relationship between deuterium and oxygen-18 delta values in leaf water.Chemical Geology:Isotope Geoscience Section,1985,58(1/2):145-156.
[57]Cernusak L A,Pate J S,Farquhar G D.Diurnal variation in the stable isotope composition of water and dry matter in fruiting Lupinus angustifolius under field conditions.Plant,Cell and Environment,2002,25(7):893-907.
[58]Yakir D,DeNiro M J,Gat J R.Natural deuterium and oxygen-18 enrichment in leaf water of cotton plants grown under wet and dry conditions:evidence for water compartmentation and its dynamics.Plant,Cell and Environment,1990,13(1):49-56.
[59]Gat J R,Yakir D,Goodfriend G,Fritz P,Trimborn P,Lipp J,Gev I,Adar E,Waisel Y.Stable isotope composition of water in desert plants.Plant and Soil,2007,298(1/2):31-45.
[60]Cooper L W,DeNiro M J,Keeley J E.The relationship between stable oxygen and hydrogen isotope ratios of water in astomatal plants//Taylor H P,O'Neil J R,Kaplan I R,eds.Stable Isotope Geochemistry:A Tribute to Sam Epstein.San Antonio:The Geochemical Society,Special Publication,1991:247-255.
[61]Cuntz M,Ogée J,Farquhar G D,Peylin P,Cernusak L A.Modelling advection and diffusion of water isotopologues in leaves.Plant,Cell and Environment,2007,30(8):892-909.
[62]Gat J R,Bowser C.The heavy isotope enrichment of water in coupled evaporative systems//Taylor H P,O'Neil J R,Kaplan I R,eds.Stable Isotope Geochemistry:A Tribute to Samuel Epstein.San Antonio:The Geochemical Society,Special Publication,1991:159-168.
[63]Farquhar G D,Lloyd J.Carbon and oxygen isotope effects in the exchange of carbon dioxide between terrestrial plants and the atmosphere//Ehleringer J R,Hall A E,Farquhar G D,eds.Stable Isotopes and Plant Carbon-Water Relations.San Diego:Academic Press,1993:47-70.
[64]Ogée J,Cuntz M,Peylin P,Bariac T.Non-steady-state,non-uniform transpiration rate and leaf anatomy effects on the progressive stable isotope enrichment of leaf water along monocot leaves.Plant,Cell and Environment,2007,30(4):367-387.
[65]Farquhar G D,Cernusak L A.On the isotopic composition of leaf water in the non-steady state.Functional Plant Biology,2005,32(4):293-303.
[66]Shu Y,Feng X,Posmentier E S,Sonder L J,Faiia A M,Yakir D.Isotopic studies of leaf water.Part 1:A physically based two-dimensional model for pine needles.Geochimica et Cosmochimica Acta,2008,72(21):5175-5188.
[67]Kodama N,Cousins A,Tu K P,Barbour M M.Spatial variation in photosynthetic CO2,carbon and oxygen isotope discrimination along leaves of the monocot triticale(Triticum × Secale)relates to mesophyll conductance and the Péclet effect.Plant,Cell and Environment,2011,34(9):1548-1562.
[68]West A G,Goldsmith G R,Brooks P D,Dawson T E.Discrepancies between isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters.Rapid Communications in Mass Spectrometry,2010,24(14):1948-1954.
[69]West A G,Goldsmith G R,Matimati I,Dawson T E.Spectral analysis software improves confidence in plant and soil water stable isotope analyses performed by isotope ratio infrared spectroscopy(IRIS).Rapid Communications in Mass Spectrometry,2011,25(16):2268-2274.
[70]Zhao L,Xiao H,Zhou J,Wang L,Cheng G,Zhou M,Yin L,McCabe M F.Detailed assessment of isotope ratio infrared spectroscopy and isotope ratio mass spectrometry for the stable isotope analysis of plant and soil waters.Rapid Communications in Mass Spectrometry,2011,25(20):3071-3082.
参考文献:
[21]吴绍洪,潘韬,戴尔阜.植物稳定同位素研究进展与展望.地理科学进展,2006,25(3):1-11.
[22]林光辉.稳定同位素生态学:先进技术推动的生态学新分支.植物生态学报,2010,34(2):119-122.
[24]周辰昕,孙自永,余绍文.黑河中游临泽地区沙丘植物水分来源的D、18O同位素示踪.地质科技情报,2011,30(5):103-109.
[25]孙守家,孟平,张劲松,黄辉,万贤崇.华北石质山区核桃-绿豆复合系统氘同位素变化及其水分利用.生态学报,2010,30(14):3717-3726.
[27]袁国富,张娜,孙晓敏,温学发,张世春.利用原位连续测定水汽δ18O值和Keeling Plot方法区分麦田蒸散组分.植物生态学报,2010,34(2):170-178.
[28]温学发,张世春,孙晓敏,于贵瑞.叶片水H182O富集的研究进展.植物生态学报,2008,32(4):961-964.
[30]张世春.大气水汽δ18O和δD原位连续观测及其应用研究[D].北京:中国科学院研究生院(地理科学与资源研究所),2010.
[49]李倩,谭雪莲.旱地植物蒸腾作用研究进展.甘肃农业科技,2006,12(10):18-20.
