克氏原螯虾幼体及亚成体对中华鳖的趋避行为
2013-07-05王陈路覃简萍
王陈路,覃简萍,黄 成
南京大学生命科学学院,江苏南京 210093
克氏原螯虾Procambarus clarkii Girard由于其食性广、生长快、生命周期相对较短、增殖速度快及后代多等特质,已成功入侵澳大利亚和南极洲以外的地方(Gherardi,2006)。克氏原螯虾相比于土著螯虾,捕食效率更高,会严重破坏两栖动物的卵和鱼虾的幼苗,对当地的生态系统造成严重的威胁(Renai&Gherardi,2004)。克氏原螯虾的捕食者主要为哺乳动物、鸟类和一部分鱼,如水貂Neovison vison Schreber、苍鹭 Ardea cinerea L.、河鲈 Perca fluviatilis L.、欧洲鳗鲡 Anguilla anguilla L.等(Blake &Hart,1995;Geiger et al.,2005)。
在种间关系中,捕食是一种常见的自然现象,对生态系统和生物多样性的稳定有着非常重要的作用,也是利用天敌进行生物防治的生物学基础。江舒等(2007)提出,利用土著天敌捕食克氏原螯虾,可以弥补化学防治的缺陷,不会对当地的生态系统造成危害。中华鳖Trionyx sinensis Wiegmann又称甲鱼,是我国普遍分布的具有重要价值的土著水生动物,主要以螺类、虾蟹、鱼类以及动物尸体等动物性饵料为食(杨振才等,1999)。笔者在初步养殖试验中发现甲鱼会捕食螯虾,但螯虾对甲鱼的反应尚不明确。因此,本试验以甲鱼的气味作为捕食者信号源,研究螯虾对甲鱼的回避行为,以期为克氏原螯虾的生物防治提供参考。
1 材料与方法
1.1 材料
试验时间为2012年4~6月和2013年5月,试验动物均来自江苏省盱眙县满江红龙虾养殖基地。幼虾体长为2~3 mm,可以离开母体自由活动,但仍常回到母体腹部。亚成体螯虾附肢健全,活力正常,接近性成熟从未交配,体质量(9.58±3.68)g;每只亚成体螯虾用直径约18 cm网笼单养驯化一星期,使其适应室内环境;第1天不喂食,第2天开始用幼蟹混合饲料喂养。按照目前的分类,幼虾属于虾苗发育阶段,而亚成体螯虾属于成虾阶段(王克行,1997)。甲鱼每只体质量(213.38 ±6.83)g,室内适应驯养一星期。
试验所用水迷宫示意图见图1。将甲鱼置入直径约35 cm的网笼,与亚成体螯虾隔离混养2 d即得有经验的亚成体螯虾,混养水箱设置见图2。

图1 水迷宫示意图Fig.1 Schematic diagram of the"Y"-maze

图2 甲鱼螯虾混养示意图Fig.2 Schematic diagram of mixed culture of crayfishes and the soft-shelled turtle
1.2 幼虾对甲鱼的趋避性试验
以60只幼虾为1组。“Y”形水迷宫两臂入口及原点出口处均用网隔隔开,试验时向迷宫内注入6~7 cm水,漫过甲鱼,两臂圆槽的一侧放置1只甲鱼,用网罩罩住圆槽以防止试验过程中甲鱼逃跑,另一侧为空白。在室内全遮光环境中,将1组幼虾放入原点,静置5 min后取出网隔开始计时,30 min后结束试验,用自制网隔(使用尼龙绳网包裹铁丝网,确保幼虾无法透过)插在水迷宫两臂口(图1所示黑白交界处),计数左右两臂(即图1所示黑色区域)的虾,总和即为外出的虾。捞出幼虾,清理水迷宫,用吹风机将水迷宫吹干,以消除试验及换水过程中气味扩散的影响,然后进行下一组试验。每天9组试验,连续进行4 d得到36组数据,第1天的9组为无经验组,后面的27组为有经验组。以水迷宫左右两端均空白作为对照组,重复10次。
1.3 亚成体虾对甲鱼的趋避性试验
以单只螯虾为1组,每个处理设30组重复,设无经验组和有经验组。在室内全遮光环境中,使用USB DVR BOX(USB2.0)红外视频拍摄,摄像头为OS-20GB。试验时向“Y”形迷宫内注入6~7 cm水,漫过甲鱼,两臂圆槽内一侧放置1只甲鱼,用双层铁丝网隔开,用网罩罩住圆槽以防止甲鱼逃跑,另一侧为空白。将螯虾放入原点,静置5 min后取出铁丝网隔开始拍摄,拍摄时间为15 min。完成后换水,清理水迷宫,将水迷宫用开水烫洗一遍,以消除气味扩散的影响,然后进行下一组试验。以水迷宫左右两端均空白作为对照组,记录5 min内留在原点和选择外出(即左右两臂)的螯虾数量。
1.4 数据处理
根据1.3拍摄的视频,统计螯虾的首次选择(选择甲鱼还是空白)、累计次数(选择甲鱼或空白多少次)及时长(选择甲鱼或空白的时间)。用SPSS 18.0和EXECL软件对试验数据进行统计分析。对螯虾亚成体的外倾性、首次选择、累计次数用二项分布进行检验;对无经验螯虾外倾性和对照组做百分数差数检验;对幼虾选择甲鱼和空白的只数以及亚成体虾选择甲鱼时间和空白时间做t-检验。
2 结果与分析
2.1 幼虾的趋避性
由表1得:水迷宫左右两端均为空白时,外出选择的幼虾数量显著高于未选,外倾性很强;当一端有甲鱼信息源时,无经验和有经验的幼虾外出选择的数量都显著低于未选,变为极显著的内倾性;且信息源试验中外出选择的幼虾数量极显著低于空白对照(P=0.003 <0.01)。
由表2得:水迷宫左右两端均为空白时,螯虾对左右的选择差异不显著;当一端有甲鱼信息源时,幼虾无论是否有经验,选择甲鱼的数量均显著低于选择空白的数量;且总体上选择甲鱼的幼虾数量也极显著低于选空白的数量。
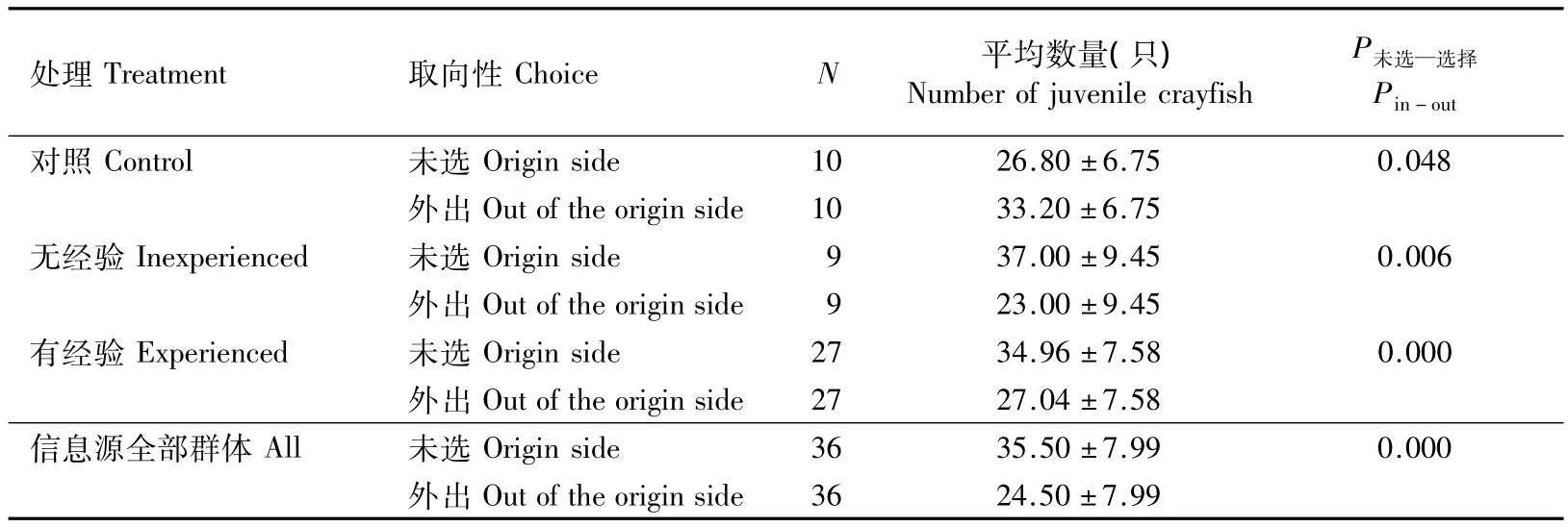
表1 幼虾外倾性结果的比较Table 1 T-test of the average number of juvenile P.clarkii in the origin side or out of the side

表2 幼虾对甲鱼选择结果的比较Table 2 T-test of the average number of juvenile P.clarkii's choice to the turtle or the blank,the same area where there is no turtle
2.2 亚成体虾的趋避性
由表3得:空白对照中,螯虾外出数量极显著高于未选,即外倾性极强,已做出选择的螯虾无左右偏向性(P=0.711>0.05)。有甲鱼信息源的试验中,无经验组螯虾外出数量与未选差异不显著,有经验组螯虾外出数量显著高于未选,即仍具有极强的外倾性。经过百分数差数检验,无经验组的外倾性极显著低于空白对照(P=0.001 <0.01),而有经验组与空白对照差异不显著(P>0.05)。
空白和甲鱼之间的首选及累计选择次数差异均不显著(表3)。此外,无经验组对非甲鱼(空白+未选)的选择显著大于甲鱼,有经验组则差异不显著。

表3 亚成体虾首选组数及累计次数的统计Table 3 First choice and frequency of the subadults
由图3可得:有经验组和无经验组在空白、甲鱼及外出时长的比较上,有经验组均大于无经验组,但差异不显著;t-检验结果表明,有经验组和无经验组对空白、甲鱼选择的累计时间差异均不显著。

图3 亚成体虾对甲鱼、空白的选择时长及外出时间的比较Fig.3 The time length of crayfish's choice to turtle,blank and all
3 讨论
3.1 化学信号在克氏原螯虾反捕食行为中的作用
螯虾识别捕食者的方法有很多,如视觉、化学感受、机械碰触等(Blake&Hart,1995)。陈楠生和孙海宝(1992)研究发现,甲壳动物主要靠体表的化学感受器感知周围环境情况,从而做出捕食、躲避敌害、觅偶交配等行为;Gherardi et al.(2011)也证明,克氏原螯虾可以通过化学通讯感受到水环境中捕食者的化学信号及其他引起螯虾反捕食行为的信号。本次试验在暗室中进行,采用红外装置拍摄,可以排除视觉信号的可能性,认为螯虾主要靠化学感受器识别捕食者;但不能排除甲鱼发出能被螯虾感知的非光学物理信号(如震动、电场等)的可能性,这有待进一步探究。
3.2 克氏原螯虾幼体和亚成体对捕食者趋避性的比较
Acquistapace et al.(2003)的试验证明,克氏原螯虾可以通过气味识别捕食者,面对捕食者时会降低取食行为和自身的活动性,降低自身对捕食者的暴露机会。本试验中,当水迷宫一端有甲鱼时,幼虾由显著外倾变为极显著内倾,亚成体外倾性也显著下降;同时,选择空白的幼虾或亚成体虾数量显著高于选择甲鱼的数量。这说明2个发育阶段的螯虾对甲鱼均有回避性。
Gherardi et al.(2011)以4种鱼作为克氏原螯虾成体的捕食者进行研究,认为螯虾可以选择性地躲避捕食能力强的物种,而对在野外长期共存的物种不表现出回避行为。本试验中,有经验组亚成体虾具有极强的外倾性,且不回避甲鱼。这可能是因为亚成体与甲鱼隔养2 d中一直未受伤害,且亚成体螯虾外壳坚硬,面对甲鱼有一定的抵抗能力,这使得螯虾通过习惯化学习(尚玉昌,2005),产生特异刺激性疲劳,避免了无谓的能量消耗,表现出对共存过的甲鱼的适应性。而有经验的幼虾具有极强的内倾性及对甲鱼的回避性。这是由于幼虾面对捕食者时没有任何的抵抗力,表现出比亚成体虾更加敏感的识别及回避的能力;此外,还与幼虾和甲鱼接触时间短暂有关。
陈楠生,孙海宝.1992.甲壳动物化学感觉研究进展.海洋与湖沼,23(3):334-342.
江舒,庞璐,黄成.2007.外来种克氏原螯虾的危害及其防治.生物学通报,42(5):15-16.
尚玉昌.2005.动物的习惯化学习行为.生物学通报,40(10):9-11.
杨振才,牛翠娟,孙儒泳.1999.中华鳖生物学研究进展.动物学杂志,34(6):41-44.
Acquistapace P,Hazlett B A and Gherardi F.2003.Unsuccessful predation and learning of predator cues by crayfish.Journal of Crustacean Biology,23:364 -370.
Blake M A and Hart P J B.1995.The vulnerability of juvenile signal crayfish to perch and eel predation.Freshwater Biology,33:233-244.
Geiger W,Alcorlo P,Baltanas A and Montes C.2005.Impact of an introduced Crustacean on the trophic webs of Mediterranean wetlands.Biological Invasions,7:49 -73.
Gherardi F.2006.Crayfish invading Europe:the case study of Procambarus clarkii.Marine and Freshwater Behaviour and Physiology,39:3,175-191.
Gherardi F,Mavuti K M,Tricarico E and Harper D M.2011.The smell of danger:chemical recognition of fish predators by the invasive crayfish Procambarus clarkii.Freshwater Biology,56:1567 -1578.
Renai B and Gherardi F.2004.Predatory efficiency of crayfish:comparison between indigenous and non-indigenous species.Journal of Crustacean Biology,18:120 -127.
