大豆耐热机制的自然积累*
2013-07-04顾和平袁星星陈华涛崔晓艳张红梅
顾和平,袁星星,陈 新,陈华涛,崔晓艳,张红梅
(江苏省农业科学院蔬菜研究所,南京 210014)
长江中下游地区和中国南方夏大豆区,大豆生长中后期常受高温逆境的威胁,严重影响大豆的产量和品质[1],2003年7月底8月初和2010年8月的连续高温,不仅给上述地区的水稻带来较大面积的严重减产[2-3],而且严重影响了大豆的产量。随着人类社会城市化步伐的不断加快,石化燃料的大量使用,地球上的温室效应日益明显,据估计到2100年,地球的平均温度还将上升1.8~4.0℃[4],高温作物遭受强高温胁迫的危险会越来越大。适当迟播,可以在一定程度上规避大豆生长中后期的高温危险,培育耐热品种也是解决大豆高产的一种方法。作物的自然驯化和获得性遗传也启迪我们,综合性状较为优良、但是耐热性较差的品种,经过多年的高温锻炼和获得性遗传,大豆的耐热性可能会得到明显改良。生物在个体发育中,当环境发生改变时,往往会发生相应的变异,相应于自然环境的变异就是适应,相应于人工环境的变异就是驯化,这些变异性状在生物学中称之为获得性状(Acquired characteristics)[5],1998年9月8日,Cairns等在英国的《自然》杂志上发表了《突变的起源》,介绍了乳糖诱导能利用乳糖的突变,认为事实上存在适应性突变(Adaptive mutation)及定向突变(Directedmatution),此后不少学者发表了适应性突变的论文,证明适应性突变不是偶然的个别现象,而是带有一定的普遍性[6]。1994年,Foster和Rosenberg等通过对大肠杆菌FC40的研究指出,细菌在环境压力下,可以主动改变遗传结构,通过重组新基因来应付外部环境的改变[7]。也有报道,长期的高温逆境会引起生物性状的改变,并且这种改变会遗传给后代[8],大面积生产遇到的高温严重减产问题经常提醒我们,很有必要对大豆的耐热性进行认真研究。制定大豆高温驯化的研究方案,使供试材料在完全模拟自然的条件下接受高温锻炼,研究高温驯化提高大豆耐热性的实际效果。明确连续高温栽培对大豆、乃至大多数农作物的抗热性影响。耐热性较差、而其他性状优良的作物新品种,在经过一段时间的适应性种植后,其耐热性和产量稳定性会有所改善。许多农家品种的耐热性优于新品种,长期的高温驯化使这些作物产生了获得性遗传可能是重要原因之一。建议新品种的更替速度不宜过快。
1 材料与方法
1.1 试验材料
1988—1989年,选用江苏省不同年代育成的代表性品种共50份,在塑料大棚内进行大豆品种耐热性筛选,以露天栽培为对照,以结荚率和籽粒产量的下降情况作为耐热性选择的主要指标。选出耐热性不同的大豆品种共6个,它们是耐热性较强品种岔路口1号、金大332,耐热性中等品种苏丰和58-161,耐热性较差品种苏系五号和苏豆1号。1990年,将上述材料全部繁殖1次,然后随机分成2份,1份用铝箔纸袋密封后放入-18℃的冷库,于2000和2010年各种植1次,以便更新种子生活力。2000和2010年的播种材料分别来自于1990和2000年收获的种子的随机样本。另1份每2年种植1次,连续种植10次,每次播种所用材料均为前一种植年份收获的种子的随机样本。每年均在6月10—20日播种,开花时间稳定在7月底到8月20日左右,让这些品种接受高温逆境的自然考验,并在后来检验其耐热性机制的自然积累程度。2010年,将连续种植过10次的6个品种编号为XH1(岔路口1号)、XH2(金大332)、XH3(苏丰)、XH4(58-161)、XH5(苏系5号)、XH6(苏豆1号)。
将存放在冷库,仅种植过2次的6个大豆品种编号为PT1(岔路口1号)、PT2(金大332)、PT3(苏丰)、PT4(58-161)、PT5(苏系5号)、PT6(苏豆1号)。
1.2 试验方法
2011年在南京六合江苏省农科院实验基地,采用4个长40m、宽8m的大棚种植上述材料。大棚内试验采用随机区组设计,小区行长4m,4行区,行距0.5m,穴距0.2m,留苗1株,重复3次。2个大棚作高温处理,即在大豆播种后30 d开始,将大棚顶部盖上塑料薄膜,使得大棚内温度白天控制在37℃,夜间控制在30℃,温度的控制依靠开关大棚左右两侧的塑料薄膜和预先装备的尼龙网纱,温度不够时,密封塑料薄膜。另外2个大棚一直不加盖塑料薄膜,大豆完全生长在自然环境下,作为对照。
1.3 统计方法
大豆成熟时,收获各小区的全部植株,并称取每个小区的实际干籽粒产量。收获前,在每个小区的中间行连续随机取样10株进行室内考种,以3个重复,共30个植株的平均值代表该品种相关农艺性状的表征值。考种后单株籽粒产量称重后并入相应小区。各处理、各品种之间农艺性状和产量的差异显著性以新复极差分析法表示。
2 结果与分析
2.1 高温栽培下耐热驯化品种和未驯化品种的小区籽粒产量表现
2.1.1 高温胁迫栽培对大豆籽粒产量的影响 由表1和表2可见,试验环境(高温胁迫栽培和常温栽培)之间的差异达极显著水平,F=13939,说明采用塑料薄膜大棚加热,实行白天(8:00~18:00)37℃、夜间(18:00~8:00)30℃的高温栽培,对检验耐热驯化品种的耐热性积累响应是有效的。
高温胁迫对全部参试材料具有显著的负向效应。综合计算,高温胁迫栽培下的大豆产量平均为1 303 g,比常温栽培的产量2 047 g下降了57.10%,达极显著水平。分类计算,经过耐热驯化的大豆产量下降了33.06%,未经耐热驯化的大豆品种的籽粒产量下降了95.60%。

表1 高温栽培下驯化品种与未驯化品种的产量比较
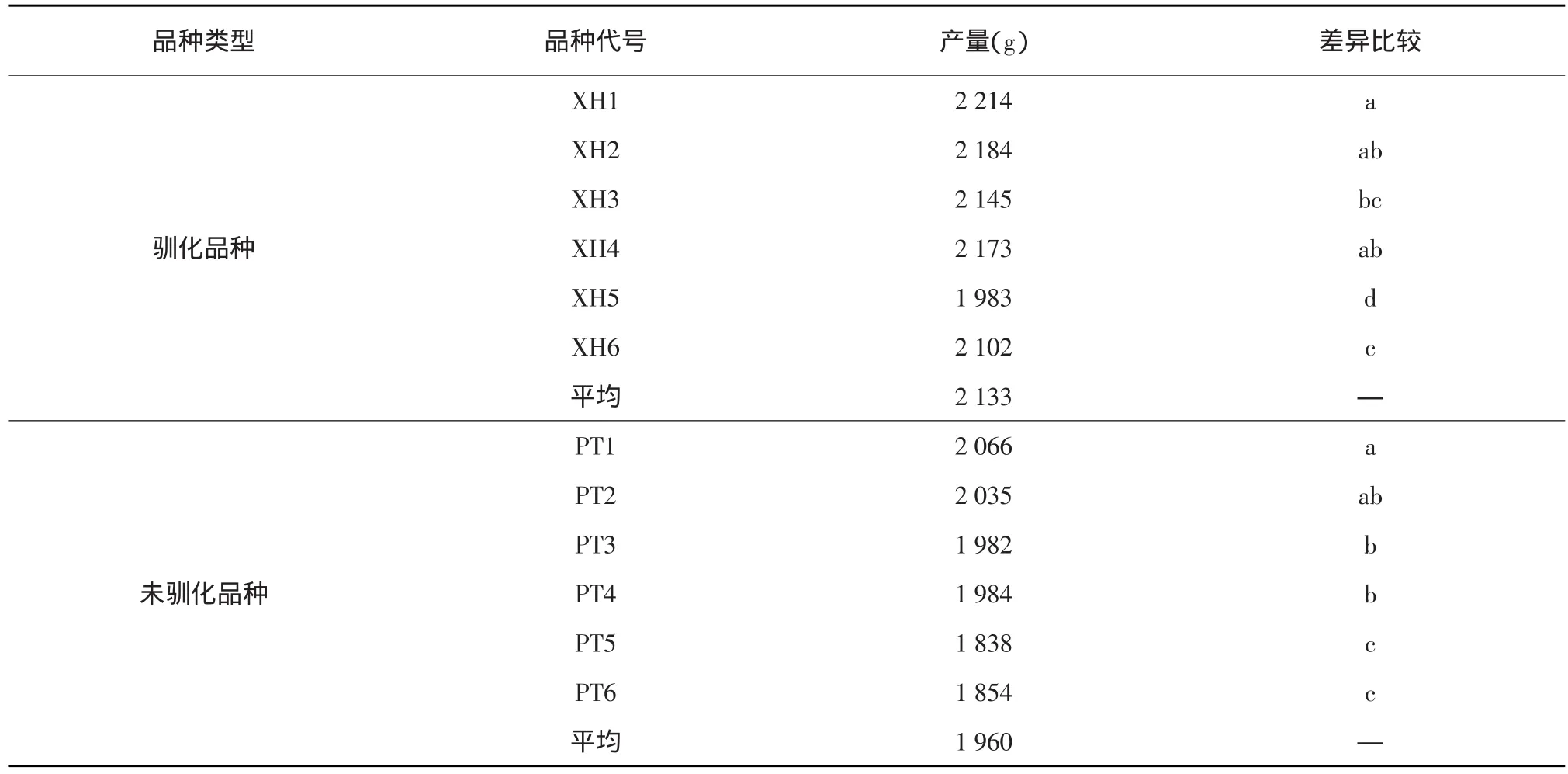
表2 常温栽培下驯化品种与未驯化品种的产量比较
2.1.2 高温栽培下驯化品种和未驯化品种的产量表现 6个品种的整体抗热性增强。在高温逆境下,经过驯化的6个品种,其小区平均籽粒产量是1 603 g,未经驯化的品种,平均产量是1 002 g,驯化品种增产了59.98%。在常温栽培环境下,高温驯化品种的平均产量是2 133 g,比未驯化品种的1 960 g增产8.83%,达显著水平(见表1和表2)。
基础耐热水平不同的品种,经过耐热驯化后,耐热性提高的程度有很大差异。本试验在高温和常温栽培下,比较了基础耐热水平不同的品种,经过10次高温种植后,与未驯化品种相比,大豆籽粒产量的差异,以产量的高低间接代表高温驯化后的耐热机制积累情况,结果表明,基础耐热性较强的品种,高温驯化后,耐热性增加了40.50%;基础水平中等的品种,经过高温驯化后,耐热性增强了47.44%;基础水平较差的品种,经过耐热驯化后,耐热性增强了114.25%。因此,我们初步认为,高温驯化可以显著提高大豆的耐热性;基础耐热性水平不同的品种对高温驯化的响应也不同,基础耐热性较强的品种,高温驯化后,耐热性提高的速度较慢,基础耐热性较差的品种,高温驯化后,耐热性增强的效果更好。
2.2 高温胁迫栽培对耐热驯化品种和未驯化品种农艺性状的影响
2.2.1 高温胁迫栽培对品种农艺性状的影响 高温胁迫对全部参试品种的农艺性状影响为,单株平均产量下降了35.82%,为16.39 g;单株荚数平均下降了30.11%,为40.29个;每荚粒数平均下降了12.40%,为1.74粒;百粒重上升了1.4%,达到22.73 g(见表3)。
2.2.2 高温栽培对耐热驯化品种和未驯化品种农艺性状的影响 由表3和表4分析可见,高温栽培对耐热驯化品种和未驯化品种农艺性状均有不同程度的影响。
对单株产量的影响:驯化品种比未驯化品种增产62.66%,达到20.30 g,未驯化品种平均是12.48 g。基础耐热性强的品种,单株产量比未驯化品种增加46.03%,达到23.00 g,未驯化品种是15.75 g;基础耐热水平中等的品种,单株产量增加了49.61%,达到19.15 g,未驯化品种是12.80 g;基础水平差的品种,单株产量增加了111.79%,达到18.85 g,未驯化品种是8.90 g。

表3 高温栽培下驯化品种与未驯化品种的产量性状比较
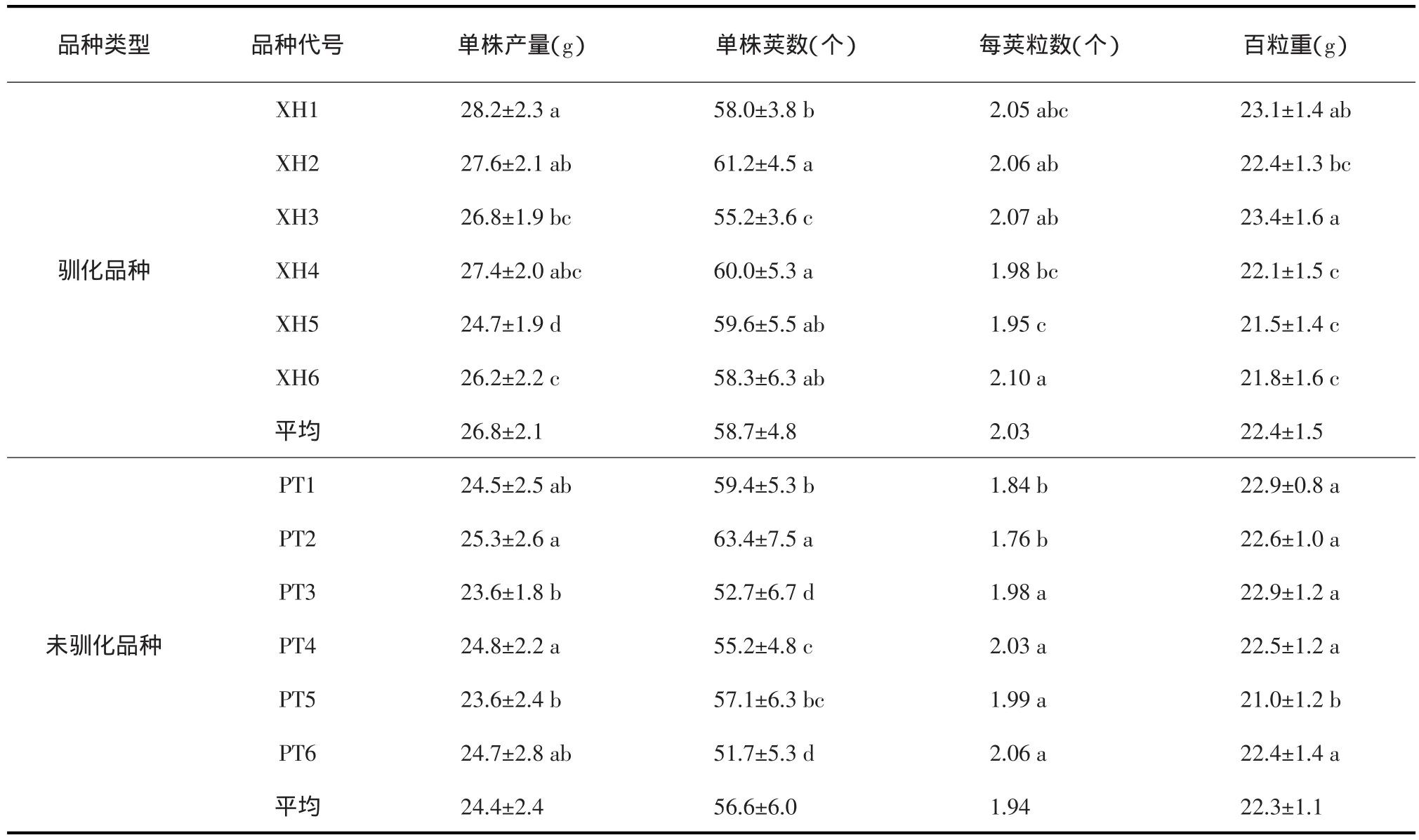
表4 常温栽培下驯化品种与未驯化品种的产量性状比较
对单株荚数的影响:驯化品种增加了43.70%,达到41.8个,未经驯化的品种平均是30.07个。基础耐热性强的品种,经过驯化荚数提高了20.59%,达到49.5个,未驯化品种,平均是41.05个;基础耐热性中等品种,驯化后荚数提高了34.11%,达到43.05个,未驯化品种平均32.1个;基础耐热性差的品种,驯化后荚数提高了91.94%,达到50个,未驯化品种是26.05个。
对每荚粒数的影响:驯化品种每荚粒数提高了13.37%,达到1.85粒,未经高温驯化的品种平均是1.63粒。基础耐热性强的品种,经过驯化后,每荚粒数提高了20.61%,达到1.99粒,未驯化品种是1.65粒;基础耐热性中等的品种,经过高温驯化后,提高了2.01%,达到1.78粒,未驯化品种是1.74粒;基础耐热性差的品种,驯化后提高了18.54%,达到1.79粒,未驯化品种是1.51粒。
对百粒重的影响:6个品种平均驯化后百粒重下降了2.74%,达到了22.45 g,未驯化品种为23.08 g。基础耐热性强的品种,驯化后百粒重下降了2.92%,为23.25 g,未驯化品种为23.95 g;基础耐热性中等品种,驯化后增加了1.08%,为23.40 g,未驯化品种为23.15 g;基础耐热性差的品种,驯化后下降了6.55%,达到20.70 g,未驯化品种为22.15 g。
综上所述,在高温胁迫栽培环境下,耐热驯化品种比未经驯化品种具有更好的抗逆境能力,在全部参试品种的水平上,单株产量、单株荚数和每荚粒数分别增加62.89%、43.70%和13.37%。百粒重则下降了2.74%。基础耐热性强的品种,在单株产量、单株荚数和每荚粒数上分别增加了46.03%、20.59%和20.61%。基础耐热性中等的品种,相应增加了49.61%、34.11%和2.01%;基础耐热性差的品种,则相应增加了111.79%、91.79%和18.54%。说明基础耐热性不同的品种,高温驯化的效果具有很大差异,越是基础耐性差的材料,高温驯化的效果就越好。
3 讨论
3.1 高温逆境下,花器是最敏感的受害部位
大量研究表明,在高温逆境下,作物的雄性花器是最敏感的受害部位。汤日圣等2006年用4个水稻品种进行高温逆境处理[9],郭晶心等2010年对不同品种水稻进行高温试验[10],李训贞等2003年对水稻花粉活力的研究都说明了这一点[11]。本次试验中,经过高温驯化的材料比未驯化材料增产69.38%,进一步分析得知,基础耐热性较强的材料,经过驯化后增产了40.50%,基础耐热性中等的材料增产了47.38%,基础耐热性较差的材料驯化后增产了114.25%。产量的增加主要来源于单株荚数的增加。通过产量构成因素可以看到,驯化品种的单株荚数平均增加了43.70%,每荚粒数仅增加了13.37%,而荚数的增加首先需要的是大豆花粉粒正常,受精条件良好。未驯化品种的产量较低,原因较多,但花粉粒生活力不强、受精环境不好肯定是重要原因之一。
3.2 高温逆境下,作物会通过多种生理反应进行自我防御
黄英金等2004年报道耐热型水稻品种在高温胁迫时,剑叶的光合速率下降幅度明显变小,胁迫解除后恢复也快,叶绿素含量、RuBpc的活性及内源多胺含量的变化趋势与光合速率相一致[12],这些均是水稻耐高温的重要生理特征。曹云英等2010年指出,高温胁迫导致水稻叶片中蛋白质的变化呈4种状态,新蛋白质的产生、一些蛋白质的表达量上调、一些蛋白质的表达被抑制、一些蛋白质的表达量下降[13]。路子显等1998年指出,高温胁迫下,大豆体内的热激蛋白含量会明显提高[14]。上述研究都从一个侧面报道了作物在高温胁迫下的自我调节和自我防御方法。实际上,作物在高温胁迫下,首先受害的还是生殖器官,因为生殖器官比之营养器官具有更大的逆境敏感性。作物在逆境下,优先考虑的是抛弃那些没有自养能力、且活力脆弱的组织和器官,这也是逆境下,生殖器官受害更重的原因之一。
3.3 大豆耐热机制的积累可能主要来源于沉默基因被不断激活
据初步研究,大豆一共有5万多个基因,数以万计的基因通过一系列非常复杂的网络型控制,决定着大豆数量性状和质量性状的形成和表达。大豆在一生中,能够表达出来的性状很可能远远低于5万个基因所能发挥的作用。很多基因在一般情况下都处于沉默状态,没有特定的环境、没有特定的刺激,这些基因将不会被激活,那些关闭的基因并不影响作物正常的生理功能。在我们的试验中,通过连续10年的高温季节种植,大豆的耐热性得到了明显的积累,和未经驯化的品种相比,产量有了显著提高,这种积累是一个逐步的和缓慢的过程。是什么造成了这种积累?我们推测,在大豆开花结荚期,每次遇到高温天气,就会有一部分耐热基因从沉默状态被激活,这种激活是不可逆的,一旦被激活,它将在后代得到稳定的遗传和表达。
大豆耐热基因的激活具有积累性和递增性,即先前被激活的基因是阈值较低的沉默基因,只有在低阈值基因全部被激活后,高能量阈值基因才会被激活。所以,经过多年的高温自然驯化,大豆耐高温的高能量阈值基因被不断激活,并不断积累和遗传下去,这样就使得大豆的耐高温能力不断加强。我们设想,在作物生长早期,让其经受多种逆境的锻炼和考验,可能会及早唤醒多种沉默基因,使其及早对多种逆境具有很好的抗性。转基因技术使人类的遗传育种事业翻开了崭新的一页,那么,通过多种刺激,唤醒多种沉默的、生物体内本身已经具备的基因也值得我们去探索。
4 结论
连续高温驯化可以显著提高大豆的耐热性,表现为在高温条件下,产量增加60%左右,在产量因素中,增加的效应依次为单株荚数大于每荚粒数大于百粒重,说明耐热性的增强反应在农艺性状上,主要是单株荚数的增加,即提高了花器官的耐热性和受孕能力。
不少农家品种的耐热性强于以产量为主要选择目标的大豆新品种,这是大豆经过高温驯化后耐热性逐步增强的佐证之一。
作物新品种的产量和抗性稳定性是作物的遗传型与所处的生态环境不断磨合、不断适应的结果,为减少作物新品种由于较大的产量选择压带来的遗传脆弱性,品种的更新不宜过于频繁。
[1]顾和平,陈新,朱成松,等.大豆耐光氧化和耐干旱种质的批量筛选鉴定[J].江苏农业学报,1999,15(1):38-41.
[2]黄义德,曹流俭,武立权,等.2003年安徽省中稻花期高温热害的调查[J].安徽农业大学学报,2004,31(4):385-388.
[3]周少川,李康活.华南一季中晚稻育种战略与研究进展[J].广东农业科学,2006(2):15-17.
[4]李敏,马均,傅泰露,等.大田生育期全程高温胁迫对杂交水稻生育后期生长发育及产量形成的影响[J].杂交水稻,2009,24(4):65-71.
[5]姚敦义,张慧娟.适应、驯化与获得性遗传[J].植物学通报,1997,14(1):19-23.
[6]韩彬,惠军,刘会强,等.探讨获得性遗传[J].新疆师范大学学报(自然科学版),2005,24(3):95-97.
[7]陈建华.反思达尔文[J].曲阜师范大学学报(自然科学版),2000,26(1):92-95.
[8]李师鹏.获得性是否可以遗传[J].生物学通报,2002,37(2):22-24.
[9]汤日圣,郑建初,张大栋,等.高温对不同水稻品种花粉活力及籽粒结实的影响[J].江苏农业学报,2006,22(4):369-373.
[10]郭晶心,曾文智,周宝津,等.开花期高温胁迫对不同水稻品种花粉萌发和结实的影响[J].华南农业学报,2010,31(2):50-54.
[11]李训贞,梁满中,周广洽,等.水稻开花时的环境条件对活力和结实的影响[J].作物学报,2002,28(3):417-420.
[12]黄英金,张红玉,郭进跃,等.水稻耐高温逼迫的生理机制及育种应用研究初报[J].科学技术与工程,2004,4(8):655-658.
[13]曹云英,段骅,王志琴,等.高温对水稻叶片蛋白质表达的影响[J].生态学报,2010,30(22):6009-6018.
[14]路子显,曲建波,傅宏仪,等.大豆热激蛋白与内源激素变化的研究[J].大豆科学,1998,17(4):318-326.
