针灸治疗肥胖的作用与分子机制
2013-06-12杜宏银张玉彬
杜宏银,张玉彬
针灸治疗肥胖的作用与分子机制
杜宏银,张玉彬
(中国药科大学生命科学与技术学院,南京 210009)
肥胖严重影响人类健康和生活质量。食品和药物管理局批准用于治疗肥胖药物奥利司他和西布曲明有严重的副反应。传统针灸法历史悠久,不仅可以用于治疗肥胖,还能用于预防和治疗肥胖引起的胰岛素抵抗和2型糖尿病。其疗效显著和无毒副反应等优点受到医学界的好评,其作用机制也引起国内外越来越多科学家的关注和研究。研究发现针灸减肥可能是通过调节下丘脑功能来控制食物的摄取和刺激一些激素如leptin,gherilin表达,发挥减肥作用。另外,针灸还可以激活外周棕色脂肪组织中b3-肾上腺素受体信号通路,增加适应性产热,达到减肥目的。文章主要综述了针灸治疗肥胖分子作用机制,为临床肥胖及相关并发症治疗提供临床理论基础。
针灸疗法;电针;肥胖症;PGC-1a;UCP1
肥胖是由多因素引起的机体能量代谢紊乱,导致脂肪组织量超过平均水平而影响人体健康的一种病理状态。随着经济的发展和人们生活方式的改变,呈逐步加重趋势,严重威胁着人类的健康。WHO的统计结果表明,目前全球有超过10亿成年人超重(BMI>25),其中至少3亿人肥胖(BMI>30)。如果不采取有效措施,预计到2015年超重人数将达15亿。肥胖症还能引发许多健康问题,不仅会增加高血压、冠心病、2型糖尿病的发病率和死亡率,还易引起呼吸系统并发症、骨关节炎以及精神方面的疾病。因此肥胖症的治疗迫在眉睫[1]。
胰岛素抵抗导致胰岛素敏感的靶组织对胰岛素应答受损,致使糖代谢和胰岛素分泌功能紊乱。胰岛素抵抗是引发2型糖尿病的重要因素,它和肥胖、2型糖尿病、多囊卵巢综合征、代谢紊乱、高血压等息息相关[2-3]。治疗肥胖的药物不多,且副反应大,价格昂贵,影响了临床使用。目前国内外很多制药公司正在积极研发新的减肥药物,可将它们分为以下3类[4]:①抑制食欲药物,通常作用于中枢神经或外周神经,通过影响食欲来减少摄食量。作用于中枢神经的有西布曲明和大麻素受体CB-1阻滞剂利莫那班,作用于外周神经系统主要有GLP-1类似物;②抑制肠道消化吸收药物,主要作用于胃肠道,减少能量吸收,主要包括奥利司他及其类似物;③增加能量消耗药物,作用于代谢过程,加快能量的消耗,主要为b3受体激动剂如SR58611A、CL316243和N25984。这些减肥药物仍处于临床前研究或临床试验阶段。
针灸在我国已运用了上千年,被广泛用于治疗肥胖症和2型糖尿病[5-6]。由于与药物治疗相比具有安全性高、副反应少、经济等优点而被人们采用。针灸治疗肥胖症的机制十分复杂,其不仅会影响下丘脑介导的饮食功能,也会引起体内代谢的变化。许多研究表明针灸不仅可以防治肥胖,还可以改善肥胖引发的高血糖、高血脂等疾病。针灸可以提高胰岛素敏感性,改善胰岛素抵抗[7]。本文主要综述了针灸治疗肥胖的分子作用机制,为临床防治肥胖及肥胖相关的代谢性疾病提供理论基础。
1 针灸通过下丘脑调节食欲减肥
在临床试验中,采用体针、耳针或者体耳结合的方法治疗肥胖的临床治疗效果都比较明显。罗树华[8]在主穴的基础上,胃肠腑热型配以曲池、合谷、公孙、内庭,脾虚湿盛型配以丰隆、阳陵泉、水分、气海,肝郁气滞型配以肝俞、膈俞、曲泉、太冲,脾肾阳虚型配以脾俞、命门、太溪,毫针刺,实证用泻法,虚证用补法,1个月为1个疗程,1个疗程后总有效率为76.7%。王燕珍[9]取神门、内分泌、三焦、肾、口、脾、肺、胃为主穴,用耳穴贴压法,治疗50例,临床治愈9例,显效10例,有效15例,无效16例,总有效率为68.0%。盛益国等[10]用电针配合耳穴治疗30例,总有效率为70.0%。Tian等[11]研究报道,将高脂诱导的单纯性肥胖大鼠固定,电针刺激其足三里、三阴交,每星期3次,连续4个星期,研究者发现伴随着食欲即摄食量下降,大鼠体重也严重下降,且2 Hz比100 Hz更有效果。
研究发现针灸减肥的机制与下丘脑中枢神经调节有关[12]。众所周知,下丘脑弓状核是食欲摄取的整合调控中枢。它具有两种神经元,这两种神经元在食物摄取上具有相反的作用。一种是能够表达抑制食欲多肽a-MSH(其前体为POMC)的神经元,另一种是能够表达促进食欲多肽NPY的神经元。下丘脑弓状核支配着表达a-MSH受体和NPY受体的二级神经元。有研究表明,2 Hz的电针处理可以诱导下丘脑弓状核中c-fos基因的表达,这暗示着2 Hz的电针处理可以对下丘脑的相关基因产生调控作用。已经有证据表明,食欲旺盛的肥胖大鼠的下丘脑弓状核中POMC的表达急剧下降,而2 Hz的电针刺激可以增加编码POMC和a-MSH的mRNA水平,但不影响NPY的水平[13]。为了更进一步观察2 Hz电针对NPY的表达影响[14],实验限制大鼠进食,每天只给1 h的进食时间,研究发现与随机喂食的大鼠相比,禁食大鼠的下丘脑弓状核中的NPY表达水平显著升高。在这种情况下,给予大鼠2 Hz的电针处理,大鼠的食物摄取量降低,并且伴随着体重降低和下丘脑中NPY的表达降低。因此,电针抑制食欲和体重下降的原因可能是a-MSH/POMC的表达升高和NPY的表达降低,如图1所示。

图1 针灸调节下丘脑功能控制饮食[15]
不仅如此,Shiraishi等[16]还发现耳针可以降低下丘脑外侧区LH(控制着进食区)神经元活性,且增加下丘脑腹侧核区VMH(控制着厌食区)神经元活性。Zhao M等[17]用电针刺激足三里和内庭穴发现,肥胖大鼠的VMH电神经活性增强。这暗示着厌食中枢的激活,由于VMH和LH都是下丘脑弓状核ARH的下游功能性中枢,ARH神经元释放的多肽可能控制着LH和VMH的神经元活性从而介导电针的效应。
最新研究发现[18]下丘脑弓状核中CART与a-MSH共存可抑制食欲,而电针刺激可以增加下丘脑弓状核中CART的多肽表达水平[19]。这暗示着电针刺激可能是通过调节下丘脑弓状核中的相关多肽从而下调食物摄取,调节能量平衡。基因组学和蛋白组学将帮助我们进一步阐明电针调控下丘脑弓状核中相关基因表达的分子机制。
2 针灸通过脂肪组织和胃肠道调节食欲减肥
针灸除了调控下丘脑中促进食欲多肽NPY和抑制食欲多肽POMC的表达,影响二级中枢,控制食物的摄取外,针灸还会影响体内一些激素如Leptin, ghrelin等表达,从而间接影响食物的摄取,最终控制体重达到减肥效果,如图1所示。
1994年,Zhang等利用定位克隆技术克隆出了肥胖基因(obese gene,ob基因),并通过定性研究发现ob基因的蛋白产物(leptin)具有抑制摄食、降低体重的作用,Leptin占据了肥胖研究的核心地位。Leptin通过其外周和神经中枢中的leptin受体发挥其功能, 1995年Ann等用ob/ob鼠做试验,研究leptin与食物摄入量的关系时发现,食物摄入减少量与leptin之间呈现剂量-效应关系。进一步的研究发现,leptin是通过直接作用于下丘脑弓状核和室旁核的受体而发挥这一作用。其次,血浆中的leptin显著升高会和脂肪组织中Leptin受体结合启动下游信号通路从而能够加速自身脂解和脂类氧化,同时伴随调节脂肪代谢和能量生成酶的基因表达量的增加。
Kim等[20]发现肥胖大鼠血浆中的leptin水平显著升高,You等[21]发现用100 Hz电针刺激足三里、内庭穴可以使得肥胖大鼠血浆中leptin显著降低,并且孙立虹等[22]发现2 Hz电针刺激大鼠曲池、足三里、丰隆、三阴交、中脘和关元穴后,下丘脑中血清leptin显著升高。这些结果暗示了电针可能促使血浆中的leptin通过血脑屏障进入下丘脑后与下丘脑leptin受体结合,促进食物的摄取。另一方面,也有研究者认为[21-22],受试者对leptin的敏感性可以看作是决定肥胖发生的更重要的因素,而非血浆中的leptin水平。因此,研究电针是否能够增强leptin的敏感性很关键,针灸对leptin敏感性的研究还需要更进一步的研究。
Ghrelin是生长激素受体内源性配体,主要由胃产生,能够增加食物摄取和体重[23]。1999年日本科学家KOJIMA等利用免疫组化的方法在小鼠和人的胃内分泌细胞中新发现了一种含有氨基酸残基的多肽,它是生长激素促分泌素受体(GHS-的内源性配体),当这种多肽与GHS-R结合后,能刺激生长素(GH)的分泌,因此将其命名为ghrelin(ghre即grow之意)。ghrelin促进摄食的作用是通过中枢食欲调节网络实现的。弓状核是下丘脑调节食物摄入的主要部位,包含促进食欲(NPY和AGRP)和减少食欲(阿片黑皮素原)的神经元。ghrelin激活位于弓状核的产生NPY/AGRP的神经元的GHS-R,促进NPY和AGRP的表达和释放,刺激食物摄入[24]。脑室内注入中和NPY、AGRP的抗体和NPYY1受体的拮抗剂,可以阻断ghrelin促进食欲的效果。有研究发现,在肥胖患者体内循环中,ghrelin和正常人相比是下降的[25],也有学者指出,在肥胖犬齿类中, ghrelin丢失了其昼夜的节律性[26],所有这些发现暗示了在肥胖者中,无论是动物还是人,ghrelin的正常功能受损。在最近的研究中,有研究者发现伴随着肥胖大鼠体重的减轻,2 Hz电针刺激可以部分修补ghrelin的昼夜节律性,但其修补ghrelin的功能机制目前还不清楚,有待于进一步研究。
3 针灸通过激活棕色脂肪组织BAT产热减肥
人和哺乳动物体中的脂肪组织分为棕色脂肪组织(BAT)和白色脂肪组织(WAT)。BAT在所有哺乳动物体中都存在,主要是幼小哺乳动物体中,而在成熟的哺乳动物体中仅有少量存在。但近年来的研究表明,户外工人和长期饮酒人群的颈动脉周围和心包存在BAT,白色脂肪组织和棕色脂肪组织作用相反,WAT主要是通过甘油三酯贮存能量,而BAT则通过产热来消耗能量从而达到减肥的作用,而这些产热主要是通过非寒栗性NTS产热来实现的。
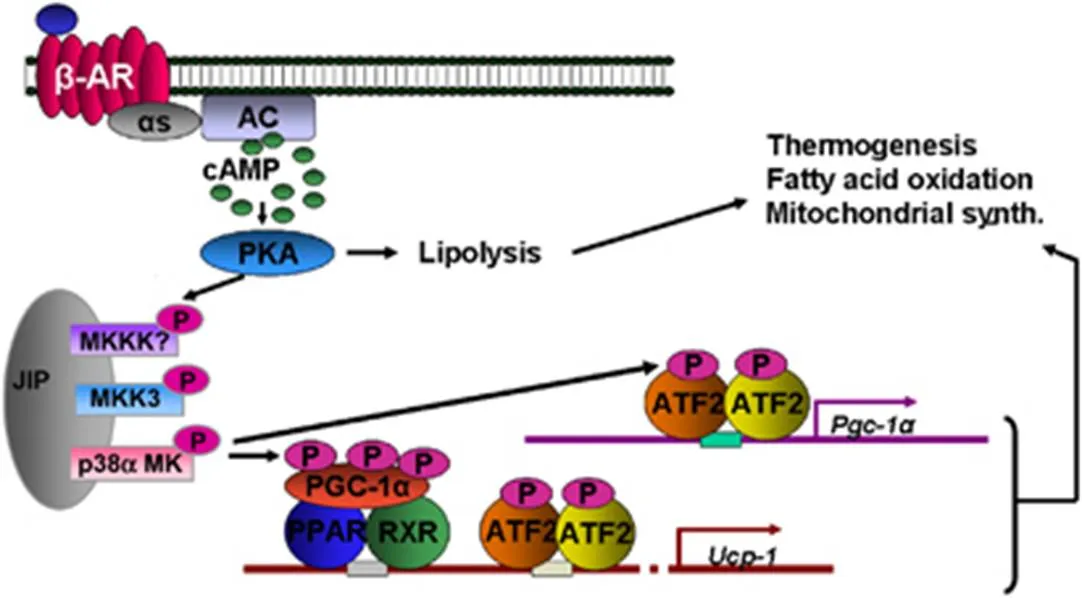
图2 棕色脂肪组织中肾上腺素能受体信号产热通路[27]
在棕色脂肪细胞中,富含线粒体。其线粒体含量远远超过白色脂肪细胞,且线粒体内膜中存在着特有的解耦联蛋白-1(uncoupling protein-1),后者可将内膜间隙的大量质子转入线粒体基质,使氧化磷酸化作用解偶联,从而降低线粒体膜电位,提高线粒体电子转运率,最终导致产热和大量化学能的消耗。UCP1的表达量和活性大小是衡量减肥作用的重要标志性蛋白,它已成为减肥治疗作用的重要靶点之一。而PGC-1a(peroxisome proliferator-activated receptor gamma coactivator-a,过氧化物酶体增殖物激活受体g辅激活受体1a)是体内能量代谢调控的重要节点蛋白,它在糖脂代谢中发挥重要作用[28]。在冷暴露条件下,去甲肾上腺素和棕色脂肪组织中b3肾上腺素能受体结合再由G蛋白偶联活化AC腺苷酸环化酶,增加细胞内的CAMP浓度增加,从而激活CAMP依赖的PKA通过肾上腺素能受体信号通路。活化的PKA还可以促进PGC-1a表达和磷酸化修饰作用增加PGC-1a的活性,从而诱导棕色脂肪细胞中UCP-1的高表达,进而脂肪分解代谢,发挥减肥作用,如图2所示。2009年,Zhang YB等[29]人报道了一种PGC-1a家族的新成员NT-PGC-1a(N-Truncated PGC-1a),NT-PGC-1a具有独特结构与功能。NT-PGC-1a的半衰期比PGC-1a长5倍,其促进肉毒碱棕榈酰转移酶Ⅰ(carnitine palmitoyltrans- ferase 1b,CPT-1b)表达活性是PGC-1a的18倍。CPT-1b是脂肪酸b氧化代谢的关键酶,表明NT-PGC-1a可以更有效促进脂肪酸氧化,将为针灸减肥而发挥重要作用,目前我们正在开展相关性研究。
近年来的研究表明,针灸减肥可能与其影响棕色脂肪组织相关基因和蛋白的表达有关。刘志诚等[30]发现,高脂诱导的肥胖大鼠的b3-AR受体基因表达降低。他们选取大鼠的足三里和内庭穴,10 Hz,1.5 V连续刺激两星期,发现肥胖大鼠b3-AR受体基因表达上升,提出了针灸可能通过促进棕色脂肪组织b3-AR受体基因的表达可能是针灸减肥的部分分子机制。不仅如此,他们还发现针灸减肥还通过加强WAT中脂肪分解。去甲肾上腺素和肾上腺素也能直接激活组织脂肪酶,促进脂肪分解,而这些激素作用是通过cAMP来实现的[31]。提示针灸提高单纯性肥胖患者cAMP水平是其产生减肥效应的重要因素。推测这可能是通过调整细胞膜上不同受体的数目或亲和力来提高cAMP水平而实现的。
4 展望
肥胖症是一种十分复杂的代谢疾病,其发生和发展涉及到了众多基因,蛋白和代谢小分子的相互作用。针灸治疗肥胖的效果是有目共睹的,但其作用机制复杂而深奥,涉及到穴位的多用性和多变性。临床上穴位选择对于治疗肥胖和2型糖尿病至关重要,不同的穴位选择可能带来不同的效应。国内一些学者对于肥胖病的临床穴位选择进行了筛选和统计,发现足三里、内庭、三阴交等穴位用到的次数频率较高,并且针对这些穴位的选择提出了一些理论。但近年来,随着现代生命科学的研究与进展,人们发现针灸治疗肥胖病的机理要通过整体水平的研究来解决,特别是基因组学和蛋白质组学的发展。这些“组学”技术不仅仅为肥胖病的领域治疗提供了新的研究方法和新技术,更为系统生物学的研究策略开辟了极大的空间。相信随着系统生物学的研究和发展,针灸治疗肥胖症的机制将会进一步得到阐明。
[1] Halford JC. Obesity drugs in clinical development[J]. Curr Opin Investig Drugs, 2006,7(4):312-318.
[2] Galluzzo A, Amato MC, Giordano C. Insulin resistance and polycystic ovary syndrome[J]. Nutr Metab CardiovascDis, 2008,18(7):511- 518.
[3] Kahn SE, Hull RL, Utzschneider KM. Mechanisms linking obesity to insulin resistance and type 2 diabetes[J]. Nature, 2006,444(7121): 840-846.
[4] 钱磊,尤启冬,廖清江,等.减肥药物最新研究进展[J].中国新药杂志,2007,16(6):437-442.
[5] Shi Y, Zhao C, Zuo XY. Clinical study on treatment of simple obesity due to spleen deficiency by acupuncture-moxibustion[J]. J Acupunct Tuina Sci, 2008,6(6):352-355.
[6] Zhang YH. Treatment of 35 cases with type 2 diabetes mellitus by acupuncture-moxibustion therapy[J]. J Acupunct Tuina Sci, 2010,8 (5):300-301.
[7] Chang SL, Lin KJ, Lin RT,. Enhanced insulin sensitivity using electroacupuncture on bilateral Zusanli acupoints (ST36) in rats[J]. Life Sci, 2006,79(10):967-971.
[8] 罗树华.针灸治疗单纯性肥胖60例临床观察[J].针灸临床杂志,2007,23(9):17-18.
[9] 王燕珍.耳穴贴压治疗单纯性肥胖50例疗效观察[J].山西中医学院学报,2011,12(1):46-47.
[10] 盛益国.针刺耳压为主对单纯性肥胖症患者腹部三围的影响[J].上海针灸杂志,2008,27(6):17.
[11] Tian DR, Li XD, Wang F,. Up-regulation of the expression of cocaine and amphetamine-regulated transcript peptide by elec- troacupuncture in the arcuate nucleus of diet-induced obese rats[J]. Neurosci Lett, 2005,383(1-2):17-21.
[12] Tian DR, Li XD, Shi YS,. Changes of hypothalamic alpha-MSH and CART peptide expression in diet-induced obese rats[J]. Peptides, 2004,25(12):2147-2153.
[13] Wang F, Tian DR, Shi YS,. Electroacupuncture modulates body weight in various energy status by differential regulation of neuropeptide expression in hypothalamus of the rat[C]. The First Chinese International Bioactive Neuropeptide Conference, 2007, Beijing.
[14] Tian N, Wang F, Tian DR,. Electroacupuncture suppresses expression of gastric ghrelin and hypothalamic NPY in chronic food restricted rats[J]. Peptides, 2006,27(9):2313-2320.
[15] Barsh GS, Schwartz MW. Genetic approaches to studying energy balance: perception and integration[J]. Nat Rev Genet, 2002,3(8): 589-600.
[16] Shiraishi T, Onoe M, Kojima T,. Effects of auricular stimulation on feeding-related hypothalamic neuronal activity in normal and obese rats[J]. Brain Res Bull, 1995,36(2):141-148.
[17] Zhao M, Liu Z, Su J. The time-effect relationship of central action in acupuncture treatment for weight reduction[J]. J Tradit Chin Med, 2000,20(1):26-29.
[18] Li HY, Hwang HW, Hu YH. Functional characterizations of cocaine- and amphetamine regulated transcript mRNA expression in rat hypothalamus[J]. Neurosci Lett, 2002,323(3):203-206.
[19] Tian DR, Li XD, Wang F,. Up-regulation of the expression of cocaine and amphetamine-regulated transcript peptide by elec- troacupuncture in the arcuate nucleus of diet-induced obese rats[J]. Neurosci letter, 2005,383(1-2):17-21.
[20] Kim SK, Lee G, Shin M,. The association of serum leptin with the reduction of food intake and body weight during electroacupuncture in rats[J]. Pharmacol Biochem Behav, 2006,83(1):145-149.
[21] You JS, Hung CC. Effect of electroacupuncture on plasma of leptin and insulin in diet-induced obese rats[J]. J Chin Med, 2005,16: 101-109.
[22] 孙立虹,梁玉磊,孙东云,等.电针刺激对雌雄两性实验性肥胖大鼠血脑leptin的影响[J].浙江中医药大学学报,2012,36(4): 430-432.
[23] Masayasu K, Kenji K. Ghrelin structure and function[J]. Physiol Rev, 2005,85(2):495-522.
[24]裴晓萌,艾华.ghrelin与摄食和肥胖的关系[J].卫生研究,2007,36(1):124-127.
[25] Mark LH, Derrick RW. Ghrelin in obesity[J]. Metab Syndr Relat Disord, 2006,4(1):37-42.
[26] Perreault M, Istrate N, Wang L,. Resistance to the orexigenic effect of ghrelin in dietary-induced obesity in mice: reversal upon weight loss[J]. Int J Obes Relat Metab Disord, 2004,28(7):879- 885.
[27] Collins S, Yehuda-Shnaidman E and Wang H. Positive and negative control of Ucp1 gene transcription and the role of β-adrenergic signaling networks[J]. Int J Obes, 201034:S28-33.
[28] Puiqserver P, Spiegelman BM. Peroxisome proliferator-activated receptor-gamma coactivator 1 alpha: Transcriptional coactivator and metabolic regulator[J]. Endocr Rev, 2003,24(1):78-90.
[29] Zhang YB, Huypens P, Adamson AW,. Alternative mRNA splicing produces a novel biologically active short isoform of PGC-1alpha[J]. J Biol Chem, 2009,284(47):32813-32826.
[30] 刘志诚,孙凤岷,赵东红,等.针刺对大鼠脂肪组织b-肾上腺素能受体基因表达的影响[J].中医杂志,2003,44(7):503-505.
[31] 刘志诚,申冬珠,周群,等.针灸对单纯性肥胖血浆环核苷酸的调整作用[J].南京中医学院学报,1991,7(3):160-163.
2012-12-20
R2-03
A
10.3969/j.issn.1005-0957.2013.08.691
1005-0957(2013)08-0691-04
国家自然科学基金项目(81072679)
杜宏银(1988 - ),男,2011级硕士生
张玉彬(1960 - ),男,教授,博士生导师
