中华蜜蜂成虫视觉系统中神经胶质细胞的类型及功能
2013-06-11李兆英
李兆英
(陕西学前师范学院生物科学与技术系,西安 710100)
神经胶质(glial cell)作为神经系统的重要组成部分之一,对神经系统的发育及功能起着重要的作用(Parker and Auld,2006)。在昆虫的神经系统尤其是视觉系统中,神经胶质的种类丰富而复杂(Stork et al.,2008)。
Ito 等(1995)将果蝇Drosophila melanogaster中枢神经系统中的神经胶质分为3种类型:表面神经胶质(surface glia),皮层神经胶质(cortex glia)和神经纤维网神经胶质(neuropil-associated glia)。Coles(1989)对蜜蜂Apis mellifera 雄蜂视网膜中神经胶质的功能进行了研究;Chotard和Salecker(2007)对的果蝇幼虫和成虫视觉系统中神经胶质的位置和形态特征进行了详细的描述;Saint Marie和Carlson(1983)描述了家蝇Musca domestica 视神经节层(lamina)内神经胶质的不同类型;Tix 等(1997) 描述了果蝇视髓层(medulla)和内、外视交叉(inner chiasmal和outer chiasmal)中神经胶质的不同类型。Hähnlein和Bicker(1997)通过组织染色和神经胶质特异核蛋白抗体免疫组织化学技术,描述了意大利蜜蜂中央复合体中神经胶质的类型以及胚后发育的特点。但是关于中华蜜蜂Apis cerana cerana 视觉系统中神经胶质的种类未见过报道。本研究通过形态解剖和组织化学等技术,对成年中华蜜蜂视觉系统中神经胶质的类型及其来源进行了比较研究,为神经胶质的种类和功能研究提供一定的理论依据。
1 材料与方法
1.1 实验材料
实验所用中华蜜蜂工蜂、雄蜂成虫购自西安大峪中华蜜蜂养殖基地。
1.2 组织化学
中华蜜蜂工蜂成虫4%多聚甲醛4℃固定过夜。梯度酒精脱水,二甲苯透明。石蜡包埋,脑部横向或水平连续切片,切片厚度5~7 μm,切片裱于涂有多聚赖氨酸的载玻片上,烘干备用。切片脱蜡后复水,苏木精-伊红染色(hematoxylin-eosin staining,HE 染色),Olympus(BX-51)显微镜观察拍照。
2 结果与分析
成年中华蜜蜂的视觉系统由复眼(compound eye,CE)和视叶(optic lobes,OL)构成。视叶又可分为3 部分,从外向内分别是视神经节层(lamina,La),视髓层(medulla,Me)和视小叶(lobula,Lo)。视小叶和视髓层之间有内神经交叉(inner chiasm,ic);视髓层和视神经节层之间有外神经交叉(outer chiasm,oc)(图1-A,B)。在视叶的各部分都有不同类型的神经胶质细胞存在。
中华蜜蜂视叶的最外层是视神经节层,在复眼基膜的下方是神经节层的有孔层(fenestrated layer),分布有表面神经胶质起源的有孔层神经胶质(fenestrated glia,fg)和类视筒神经胶质(pseudocartridge glia,pg)。在视神经节层神经元细胞体之间有卫星神经胶质(satellite glial cells,sg)。在神经节层,视网膜光感受器神经元发出的轴突被上皮神经胶质(epithelial glia,eg)包裹,分隔成束,形成视筒样结构(lamina cartridges),一直延伸穿过边缘神经胶质(marginal glia,mg),进入外视交叉(图1-C,D)。
与视神经节层相比,视觉中枢其他部分的神经胶质类群相对简单一些。视髓层和视小叶都被神经纤维网神经胶质(视髓层神经纤维网神经胶质,medulla neuropil glial cells,mng;视小叶神经纤维网神经胶质,lobula neuropil glial cells,long)所包围(图1-G),在它们周围划出边界。此外,在外视交叉和内视交叉中也有神经胶质细胞——外视交叉神经胶质(outer chiasmal ocg)(图1-E)和内视交叉神经胶质(inner chiasmal glia,icg)(图1-G),它们将神经纤维包裹成束,在视交叉中形成等距离的间隔。在外视交叉中有2种类型的神经胶质,小神经胶质(outer chiasm small glia,ocsg)和巨大神经胶质(outer chiasm giant glia,ocgg)(图1-E,F)。星形神经胶质(astrocyte glia,ag)位于视髓层和视小叶的基部(图1-H)。
3 结论与讨论
3.1 中华蜜蜂成虫视觉系统中神经胶质的类型及功能
同中枢神经系统其他部分相比,昆虫视觉系统中神经胶质的类型丰富而复杂。主要包括3 大类:表面神经胶质,皮层神经胶质和神经纤维网神经胶质,每一个类群的神经胶质根据其位置和形态又可细分为许多小的类型(Ito et al.,1995)。
在成年中华蜜蜂整个脑的周围有一层小而扁长形的神经束膜神经胶质,它们主要控制中枢神经系统的渗透性,承担血脑屏障的功能(Awasaki et al.,2008)。皮层神经胶质位于脑皮层内,它们伸出网状突起包围附近的多个神经细胞体。在视觉系统中,所有的皮层细胞都被称为卫星神经胶质。皮层神经胶质的功能可能是为神经元提供新陈代谢支持,也可能是通过空间约束来调节神经细胞体(Awasaki et al.,2008)。Coles(1989)的研究发现,在蜜蜂A.mellifera 雄蜂的视网膜中,主要的神经胶质可以调节神经细胞外的K+和Na+的动态平衡,并且还调节神经元的糖类代谢,它们的功能类似于视叶中的皮层神经胶质。
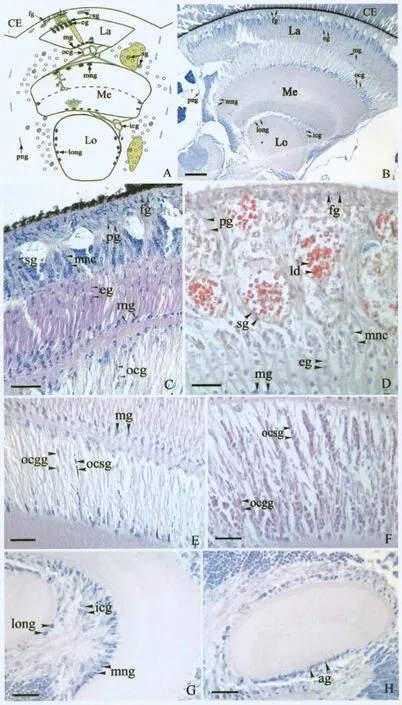
图1 中华蜜蜂成虫视觉系统的神经胶质Fig.1 Organization of glial cells in the visual system of Apis cerana cerana
神经纤维网神经胶质同轴突及轴突束相关联,它们伸出鞘状膜包裹轴突束。中华蜜蜂的神经纤维网神经胶质分为2种类型,界面神经胶质(interface glia)和星形神经胶质。界面神经胶质的突起沿着神经纤维网的外表面延伸,使神经纤维网隔离;星形神经胶质细胞的突起延伸进入神经纤维网,同突触形成联系。神经纤维网神经胶质的功能可能是使相邻的神经纤维网之间绝缘,也为神经纤维网提供内部的分隔,此外还为神经提供营养支持(Meyer et al.,1987)。
在中华蜜蜂的内、外视交叉中,有内视交叉神经胶质和外视交叉神经胶质。它们包裹轴突束,在视交叉中形成等距离的间隔。并且外视交叉神经胶质又分为2种类型—小神经胶质(outer chiasm small glia,ocsg)和巨大神经胶质(outer chiasm giant glia,ocgg)(Edwards and Meinertzhagen,2010)。Ocsg 在内外视交叉中都有,它们包裹神经纤维成束状;ocgg 仅位于外视交叉中,它们在轴突束之间排列成行。
3.2 中华蜜蜂成虫视神经节层中神经胶质的类型及功能
在中华蜜蜂成虫视觉系统的视神经节层中,有6 中不同形态学特征的神经胶质类型,包括2种表面神经胶质—有孔层神经胶质(fg)和类视筒神经胶质(pg),2种皮层神经胶质—末梢卫星神经胶质(distal satellite glia,dsg)和近端卫星神经胶质(proximal satellite glia,psg),2种神经纤维网神经胶质—上皮神经胶质(eg)和边缘神经胶质(mg)。
在中华蜜蜂成年雄蜂,有孔层神经胶质均匀的排布在视网膜基膜之下,形成了一层神经胶质细胞层,这种排列方式同家蝇M.domestica类似(Saint Marie and Carlson,1983)。但是中华蜜蜂工蜂的有孔层神经胶质排列得则较为杂乱,在基膜下形成了多层的细胞。有孔层神经胶质伸出的突起不只是包裹光感受器轴突,而且会穿过基膜进入视网膜。从有孔层神经胶质的位置来看,它们常常被归于表面神经胶质,作为血脑屏障的一部分。实际上,有孔层神经胶质具有隔膜的功能,能够为溶质的胞外扩散提供障碍(Juang and Carlson,1992)。类视筒神经胶质位于有孔层神经胶质的下面,通过细胞体的形态和位置可以识别它们。它们具有长的胞核和大的细胞体,能够完全包裹每个小眼的一束轴突。这些神经胶质类似于亚神经束膜神经胶质,它们位于视觉系统的内外交界处,构成了血脑屏障的障碍层(Saint Marie and Carlson,1983)。
卫星神经胶质是皮层神经胶质的一种,在神经节层中可分为截然不同的2种类型:末梢神经胶质,它们在神经节层皮层包裹单级神经元的细胞体;近端神经胶质,它们靠近光感受器轴突束并包绕单级神经细胞。这2种类型的神经胶质在果蝇也存在。但是因为没有特异性的标记,在蜜蜂中无法区别。在家蝇,这些细胞之间有大量的间壁连接,可能扮演了次级血脑屏障的作用(Edwards and Meinertzhagen,2010)。
上皮神经胶质属于神经纤维网神经胶质,它们延伸进入神经节层神经纤维网。上皮神经胶质具有胞核,胞体狭长,形成了独特的形态学特征。视神经节层神经细胞群是被3个一组的上皮神经胶质所包绕,形成了柱状结构,每一个神经胶质细胞同样包裹3个临近的柱状结构。上皮神经胶质的膜高度分枝,包裹神经细胞在每一个视觉筒周围形成独立的电子屏障,同时在视网膜和神经节层之间生成高电阻(Saint Marie and Carlson,1983)。边缘神经胶质覆盖在神经节层的近端表面,它们的顶点延伸突起进入神经节层神经纤维网,包围所有的轴突。这样,边缘神经胶质从基部将神经节层同其他结构分隔开(Edwards and Meinertzhagen,2010)。
References)
Awasaki T,Lai SL,Ito K,Lee T,2008.Organization and postembryonic development of glial cells in the adult central brain of Drosophila.J.Neurosci.,28:13742-13753.
Chotard C,Salecker I,2007.Glial cell development and function in the Drosophila visual system.Neuron Glia Biology,3:17-25.
Coles JA,1989.Functions of glial cells in the retina of the honeybee drone.Glia.2:1-9.
Edwards TN,Meinertzhagen IA,2010.The functional organisation of glia in the adult brain of Drosophila and other insects.Progress in Neurobiology,90:471-497.
Hähnlein I,Bicker G,1997.Glial patterning during postembryonic development of central neuropiles in the brain of the honeybee.Dev.Genes Evol.,207:29-41.
Ito K,Urban J,Technau GM,1995.Distribution,classification,and development of Drosophila glial cells in the late embryonic and early larval ventral nerve cord.Dev.Genes Evol.,204:284-307.
Juang JL,Carlson SD,1992.A blood brain barrier without tight junctions in the fly central nervous system in the early postembryonic stage.Cell Tissue Res.,270:95-103.
Meyer MR,Reddy GR,Edwards JS,1987.Immunological probes reveal spatial and developmental diversity in insect neuroglia.J.Neurosci.,7:512-521.
Parker RJ,Auld VJ,2006.Roles of glia in the Drosophila nervous system.Semin.Cell Dev.Biol.,17:66-77.
Saint Marie RL,Carlson SD,1983.The fine structure of neuroglia in the lamina ganglionaris of the housefly,Musca domestica L.J.Neurocytol.,12:213-241.
Stork T,Engelen D,Krudewig A,Silies M,Bainton RJ,Klämbt C,2008.Organization and function of the blood-brain barrier in Drosophila.J.Neurosci.,28:587-597.
Tix S,Eule E,Fischbach KF,Benzer S,1997.Glia in the chiasms and medulla of the Drosophila melanogaster optic lobes.Cell Tissue Res.,289:397-409.
