糖皮质激素诱导A549细胞11β-羟基类固醇脱氢酶1型选择性启动子的使用*
2013-06-11韩岩岩
韩岩岩
(遵义医学院公共卫生学院,贵州遵义 563099)
11β-羟基类固醇脱氢酶1型(11β-HSD1,基因名字HSD11B1)为糖皮质激素的代谢酶,它催化糖皮质激素C11位的酮基与羟基之间的氧化还原反应,使无活性的17羟11脱氢皮质酮(可的松)转化为有生物活性的皮质醇(人类),主要作用是调节局部组织糖皮质激素的浓度[1]。在啮齿类,它催化皮质酮与脱氢皮质酮之间的转化[2]。11β-HSD1的表达和活性改变可导致糖皮质激素代谢紊乱,易引发肥胖症、2型糖尿病、高血压等代谢综合征[3-5]。直到最近人们才发现,11β - HSD1基因的表达调控是由选择性启动子控制的[6],分别是位于远端的启动子1(Promoter 1,P1)和位于近端的启动子2(Promoter 2,P2)。目前关于11β-HSD1选择性启动子的研究不是很多。糖皮质激素是已知HSD11B1的调节子,可以诱导HSD11B1的表达。本文主要是探讨糖皮质激素(GC)诱导HSD11B1表达其选择性启动子的使用情况,为进一步研究GC诱导的HSD11B1表达情况和基因的转录调控模式奠定基础。
1 材料与方法
1.1 细胞培养 A549细胞(肺腺癌细胞系)培养在37℃5%二氧化碳90%湿度环境中,用DMEM培养液,添加10%的胎牛血清、青霉素(100 U/mL)、链霉素(0.1 mg/mL)和谷氨酰胺(2 mM),所有培养细胞的产品来自PAA实验室(Coelbe,德国)。
1.2 提取基因组DNA 用机械法收集单层A549细胞,然后加入1 mL细胞裂解液。基因组DNA的提取采用酚/氯仿法:悬浮液中加入1 mL的苯酚旋涡搅拌1 min。将悬浮液在室温下孵育5 min,然后13 000 r/min离心5 min。上相液体被转移到一个新的离心管,加氯仿1 mL旋涡搅拌1 min,混合物再以1 3000 r/min离心1 min(两次)。上相液体被转移到一个新的离心管,加入两个体积的100%乙醇,然后在13 000 r/min离心10 min。片状DNA颗粒用2 mL 70%乙醇洗涤,然后以13 000 r/min离心5 min。最后,DNA颗粒在室温下干燥5 min,溶于500 μL TE缓冲液。
1.3 荧光报告载体的构建 克隆11β-HSD1远端启动子 1(P1,2.173 kb)和近端启动子(P2,2.506 kb),用A549细胞基因组DNA为模板,引物(见表1)。将PCR产物克隆入pCR2.1-TOPO载体的多克隆位点,通过测序鉴定插入片段。然后将P1和P2片段再次克隆到pGL3荧光报告载体,获得目的荧光报告载体pGL3-P1和pGL3-P2。
1.4 半定量RT-PCR A549细胞用6孔板培养,传代1 d后,待细胞汇合度达90%,加入皮质醇(Cortisol,100 nM)和地塞米松(Dexamethasone,Dex,100 nM),48 h后,提取细胞总RNA用Master Pure RNA纯化试剂盒。反转录反应用2 μg总RNA,然后根据反转录试剂盒的说明操作。取1 μL反转录产物进行半定量的PCR反应,加入1 U的Phire Hotstart DNA聚合酶,1 mM的dNTP Mix,1倍的Phire反应缓冲液,0.2 μM 的引物(见表1)。PCR反应条件如下:初始变性时间98℃30 s;25个周期循环,98℃变性10 s,退火60℃15 s,延伸72℃15 s;最后一个延伸时间72℃10 min。最后,PCR产物全部用于1%凝胶电泳分离。
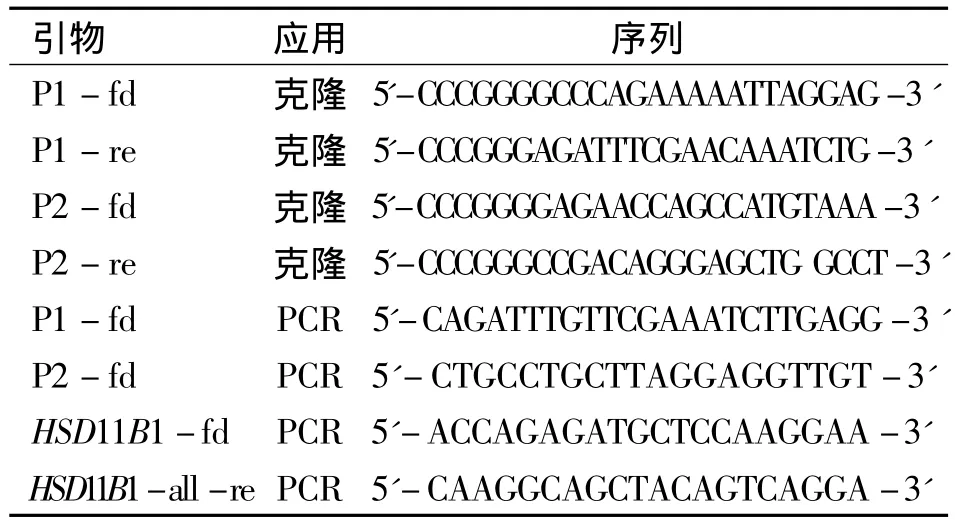
表1 实验所用引物
1.5 细胞转染 A549细胞用96孔板培养,传代1 d后,待细胞汇合度达90%,用无抗生素细胞液换液。质粒DNA(pGL3-P1和pGL3-P2,50 ng)分别用 25 μL Opti- MEM Reduced Serum Medium 稀释待用,细胞转染试剂 LipofectamineTM2000(Invitrogen GmbH,Germany)0.3 μL 用 25 μL Opti-MEM Reduced Serum Medium稀释,混合后室温放置5 min。将已稀释好的质粒DNA与稀释好细胞转染试剂混合后室温放置20 min,然后将50 μL的混合物加入每孔中。将细胞板放入37℃细胞培养箱,4 h后,将稀释好的50 μL含有糖皮质激素Cortisol(100 nM)和 Dexamethasone(Dex,100 nM)的溶液,加入细胞培养孔中,将细胞放入细胞培养箱进行孵育。
1.6 荧光报告基因分析 在细胞转染后48 h,将96孔细胞培养板移出细胞培养箱移去细胞培养液,每孔加入75 μL细胞培养液。然后,每孔加入75 μL Luciferase Reagent(Promega,Germany),将细胞板上下左右来回轻轻晃动,10 min。然后将细胞裂解物转移到96孔白色微孔板。测量firefly发光用 GENios Pro microplate reader(Tecan GmbH,Germany)。取出平板每孔加入75 μL Dual-GloTM& Glo Reagent(Promega,Germany),轻轻混合后,放置10 min。Renilla发光测量使用GENios Pro microplate reader(Tecan GmbH,Germany)。结果分析:Relative luciferase activity(%)= RLUfirefly/RLURenilla(%),其中RLU代表相对发光单位。
2 结果
2.1 GC诱导HSD11B1 mRNA的表达 糖皮质激素(GC)例如地塞米松(dexamethasone,Dex)和皮质醇(cortisol)都是非常重要的HSD11B1的调节子,可以诱导HSD11B1的表达。为了探讨GC诱导A549细胞中HSD11B1 mRNA的表达其选择性启动子的使用情况,设计特异性引物扩增来自不同启动子的转录产物(见图1)。A549细胞用地塞米松和皮质醇处理48 h后,用半定量-RT-PCR的方法检测GC诱导后HSD11B1 mRNA的表达情况。结果显示HSD11B1 mRNA表达显著增加经由选择性启动子P1和P2,但是P2要强于P1,GC主要通过介导P2诱导HSD11B1 mRNA表达(见图2)。
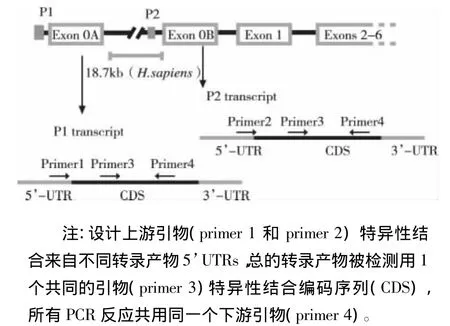
图1 设计引物检测HSD11B1选择性启动子(P1和P2)的使用情况
2.2 GC诱导荧光报告基因蛋白的表达 从A549细胞基因组中克隆11β-HSD1的两个启动子的基因序列,P1片段长为2.173 kb;P2片段长为2.506 kb(见图3)。将含有P1和P2启动子片段的荧光报告载体pGL3-P1和 pGL3-P2转染到A549细胞。经地塞米松和皮质醇处理后,检测荧光报告载体表达情况,结果显示,GC诱导的P2荧光报告载体相对荧光活性显著增加,然而P1荧光报告载体相对荧光活性没有明显变化(见图4)。

图2 半定量-RT-PCR检测HSD11B1转录产物

图3 HSD11B1选择性启动子P1和P2的PCR产物
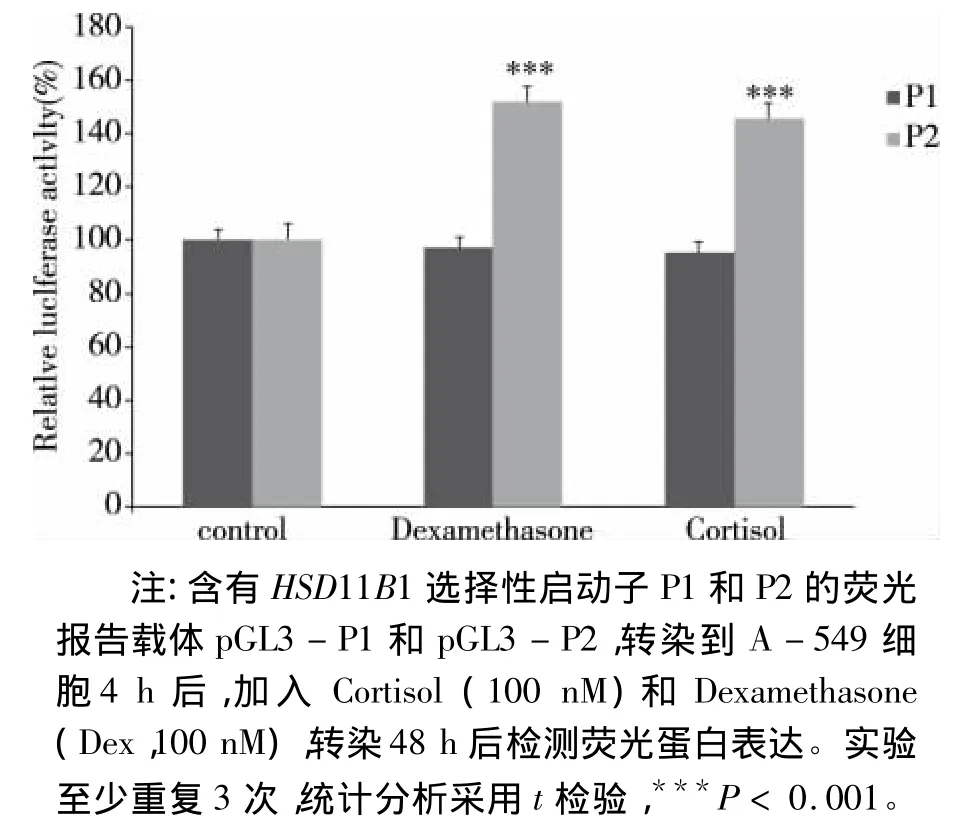
图4 GC诱导荧光报告基因的表达
3 讨论
一直以来人们对HSD11B1基因的启动子研究比较少,最近人们研究发现调控HSD11B1基因表达的启动子是由选择性启动子决定的。选择性启动子是指基因上游调控区存在的多个启动子区,每个启动子所启动的表达模式不同,这些启动子的选择性启动在基因表达中起重要的调控作用。研究发现大约有52%的人类编码基因含有2个或者多个选择性启动子,平均每个人类编码基因含有3.1个选择性启动子[7]。最近人们对选择性启动子进行深入研究,发现选择性启动子在选择性剪接基因调控中起到了重要的调控作用[8]。HSD11B1基因表达也是由选择性启动子调控的,研究发现启动子P2在大多数组织和细胞基因转录中占主导地位,包括人类肝脏、皮下脂肪组织和几种常用的细胞系如 Caco-2、C2C12 和3T3 -L1[9]。转录来自于 P1调控的主要出现在人类的肿瘤细胞,如A431和HT-29。此外,C2C12成肌细胞和3T3-L1脂肪细胞分化的过程中,HSD11B1基因转录水平增加主要来自于P2转录增加[9]。本试验应用A549细胞系是人肺腺癌细胞系,通过实验发现选择性启动子P2在肺腺癌细胞中GC诱导HSD11B1表达过程中起主导作用。
此外,本试验结果显示糖皮质激素皮质醇和地塞米松都可以诱导11β-HSD1在转录和翻译水平表达增加,其中皮质醇是11β-HSD1酶氧化还原的产物,地塞米松是皮质醇衍生物,两者匀可导致11β-HSD1酶活性增加。研究发现11β-HSD1酶活性增加,可导致机体代谢紊乱易发生糖尿病、肥胖症、高血压、胰岛素抵抗和骨质疏松等代谢疾病[10-14]。目前,糖皮质激素常用于临床疾病的治疗,糖皮质激素如皮质醇和地塞米松,临床上常用于抗炎、抗过敏、抗病毒和免疫抑制作用的治疗。短期(7 d以内)适量使用GC几乎无副作用,但是长期使用GC副作用明显,会导致物质代谢紊乱,表现为新陈代谢的变动,如血糖升高、食欲增加、体重上升、性欲减退以及极度疲劳等。长期应用糖皮质激素会导致向心性肥胖、糖尿病、高血压等疾病的发生[15-16]。综上所述,无论是糖皮质激素诱导11β-HSD1活性增加还是糖皮质激素本身长期应用对人体都有不利的影响。
本试验通过研究GC诱导HSD11B1基因表达,其选择性启动子的使用情况。结果发现,糖皮质激素诱导HSD11B1表达主要经由选择性启动子P2,该结果为今后研究HSD11B1基因的转录调控模式奠定基础。
[1]Kallberg Y,Oppermann U,H,et al.Shortchain dehydrogenases/reductases(SDRs)[J].Eur J Biochem,2002,269(18):4407 -4417.
[2]Blum A,Maser E.Enzymology and molecular biology of glucocorticoid metabolism in humans[J].Prog Nucleic Acid Res Mol Biol,2003,75:173 -216.
[3]Rask E,Olsson T,S,et al.Tissue-specific dysregulation of cortisol metabolism in human obesity[J].J Clin Endocrinol Metab,2001,86(3):1418-1421.
[4]Tomlinson J W,Stewart P M.Cortisol metabolism and the role of 11beta-hydroxysteroid dehydrogenase[J].Best Pract Res Clin Endocrinol Metab,2001,15(1):61 -78.
[5]Andrews R C,Rooyackers O,Walker B R.Effects of the 11beta-hydroxysteroid dehydrogenase inhibitor carbenoxolone on insulin sensitivity in men with type 2 diabetes[J].J Clin Endocrinol Metab,2003,88(1):285 -291.
[6]Bruley C,Lyons V,Worsley A G,et al.A novel promoter for the 11beta-hydroxysteroid dehydrogenase type 1 gene is active in lung and is C/EBPalpha independent[J].Endocrinology,2006,147(6):2879 -2885.
[7]Kimura K,wakamatsu A,Ota T,et al.Diversification of transcriptional modulation:Large-scale identification and characterization of putative alternative promoters of human genes[J].Genome Res,2006,16(1):55 -65.
[8]Hoivik E A,Witsoe S L,Bergheim I R,et al.DNA methylation of alternative promoters directs tissue specific expression of Epac2 isoforms[J].PLoS One,2013,8(7):e67925.
[9]Staab C A,Stegk J P,Haenisch S,et al.Analysis of alternative promoter usage in expression of HSD11B1 including the development of a transcript-specific quantitative real- time PCR method [J].Chem Biol Interact,2011,191(1-3):104-112.
[10]Duplomb L,Lee Y,Wang M Y,et al.Increased expression and activity of 11 beta-HSD1 in diabetic islets and prevention with troglitazone[J].Biochem Biophys Res Commum,2004,313(3):594 -599.
[11]Bays H E,Chapman R H,Grandy S.The relationship of body mass index to diabetes mellitus,hypertension and dyslipidaemia:comparison of data from two national surveys[J].Int J Clin Pract,2007,61(5):737 -747.
[12]Cooper M S,Rabbitt E H,Goddard P E,et al.Osteoblastic 11beta-hydroxysteroid dehydrogenase type 1 activity increases with age and glucocorticoid exposure[J].J Bone Miner Res,2002,17(6):979 -986.
[13]Davani B,Khan A,Hult M,et al.Type 1 11beta- hydroxysteroid dehydrogenase mediates glucocorticoid activation and insulin release in pancreatic islets[J].J Biol Chem,2000,275(45):34841 -34844.
[14]Liu J,Wang L,Zhang A,et al.Adipose tissue -targeted 11β-hydroxysteroid dehydrogenase type 1 inhibior protects against diet- induced obesity[J].Endocr J,2011,58(3):199-209.
[15]Prasad Sakamuri S S,Sukapaka M,Prathipati V K,et al.Carbenoxolone treatment ameliorated metabolic syndrome in WNTN/36 obese rats,but induced severe fat loss and glucose intolerance in lean rats[J].PLoS One,2012,7(12):50216.
[16]Chapman K E,Coutinho A E,Zhang Z,et al.Changing glucocorticoid action:11β -hydroxysteroid dehydrogenase type 1 in acute and chronic inflammation[J].J Steroid Biochem Mol Biol,2013,137:82 -92.
