乒乓球运动员动作识别时的脑电相干性分析
2013-05-23王丽岩李安民王洪彪
王丽岩,李安民,王洪彪
1 前言
运动员长期运动技能学习的外在表现之一,就是较好的运动操作绩效,而长期运动技能学习导致运动员大脑皮层变化的神经心理机制尚不清楚。目前运动员专项任务加工时的神经心理机制受到研究者的广泛关注,研究运动员认知加工神经心理的机制对于了解经验和训练对脑部神经活动的影响具有重要意义,也将有助于揭示长期运动技能学习对大脑皮层信息加工通路带来的影响。Hatfield和Hillman(2001)认为,较好的运动操作技能与运动专家完成给定任务时大脑皮层有效的加工有关[19],该观点与神经效率假说相一致。神经效率假说认为,在执行认知功能测试时,拥有良好表现的个体具有更有效的皮质功能[16]。有效的皮质功能表现在大脑皮层特定频率同步化(synchronization)振荡能量的增减,也表现在特定频率同步化的功能耦合。
近年来,脑电同步化研究主要集中在脑电信号在频域的相干性分析上。脑电的相干性分析(Electroencephalographic coherence analysis)是描述两个时间序列信号在频域内相似程度的方法[1],其在最近几年广泛应用于认知加工神 经 心 理 机 制 的 研 究 之 中[12,13,14,15]。 分 析 同 步 记 录 的 两个电极对之间具体的脑电相干性,可以用来解释皮层功能耦合的指数[10]、皮层间相互信息的交换[30]、皮 层 功 能[18]和神经通路的整合[24]。相干性基本的理论假设是,当两个皮层活动时,功能协调的脑区脑电频率显示了线性相关和高频谱的相干性。
已有研究者基于运动专家专项任务操作时“神经效率”的视角,使用相干性分析对运动员大脑皮层的功能耦合进行了探索。Haufler等(2002)对射击运动员射击时的相干性分析显示,在瞄准期间与新手相比,运动专家在高频alpha和低频beta频段内双侧大脑半球显示了整体较低的皮层活动[20]。对手枪射击运动员的研究显示,与业余运动员相比,专家运动员在低频alpha和低频beta频段左侧颞叶(T3)和中部额叶(Fz)之间显示了较低的相干性,高频alpha频段内整个颞叶与中部额叶(FZ)之间相干性低,在低频beta频段左侧颞叶(T3)与所有中线电极点之间相干性低。研究者认为,与业余射手相比,专家射手左侧颞叶与运动控制脑区之间功能耦合性低,表明了运动过程中认知参与的减少[12]。有趣的是C.DEL PERCIO等(2011)也采用专家-新手范式对手枪射击运动员的研究显示,在瞄准射击时与新手相比,运动专家在低频和高频alpha频段内顶颞叶和顶枕叶脑区展现了高相干性,在半球间不同频段顶叶也展现了高相干性[14]。而Claudio Babiloni等(2011)对高尔夫球手的研究中也显示,在成功的操作中半球内顶额叶和顶颞叶脑区间alpha频段的相干性高[9]。
运动专家大脑皮层功能耦合的研究在射击、射箭、高尔夫等运动项目中进行,不同项群的运动项目执行不同的任务时可能会导致运动员不同的大脑功能耦合和连通。因此,本研究采用专家-新手范式,应用脑电的事件相关相干性分析,对以隔网对抗性运动项目为代表的乒乓球运动员在动作识别时大脑皮层的耦合性进行研究。本研究有两个研究目的:1.探讨运动水平与脑区相干性之间的关系;2.探讨乒乓球运动员动作识别过程不同脑区之间的功能耦合。研究假设为:1.相对于乒乓球新手,乒乓球运动员在较好的操作中不同脑区展现了低相干性;2.乒乓球运动员动作识别“神经效率”的机制是涉及任务相关的注意过程和视觉运动转换的后部脑区的功能耦合。
2 方法
2.1 研究对象
本实验的参加者共为36人,其中,18名体育学院乒乓球专业的运动员(男13人,女5人),运动等级为二级,年龄19.06±2.362岁,训练年限11±2.60年;18名为乒乓球初学者(男13人,女5人),年龄20.88±1.21岁,训练年限0.88±0.18年。所有实验参加者均为右利手。实验参加前皆熟悉本研究的目的、实验要求、实验方法及流程,自愿参加本次实验并签署知情同意书,实验后予以一定的报酬。
2.2 实验材料
实验材料为由2名发球技术好的二级乒乓球运动员按实验要求发球,用摄像机拍摄其技术动作。发球旋转分为转、不转两类。摄像机为JVC高速摄像机,快门1/200,拍摄距离4.75m,镜头与地面垂直高度为1.5m。选取的发球视频片段从开始抛球动作开始,到球过中间球网,之后把所拍摄录像导出为AVI格式的视频文件。
2.3 实验仪器
E-Prime 2.0心理实验程序专业设计软件,计时精度毫秒级,用于编制实验和刺激播放程序。事件相关电位记录系统:德国Brain Products公司生产的64导事件相关电位测试系统。计算机:两台DELL计算机。一台用于运行实验程序,记录和采集行为数据,并给被试呈现刺激材料,分辨率为1 024×768,刷新频率为100Hz。另一台用于记录和采集脑电数据。
2.4 实验程序
实验在某体育学院运动心理研究中心的脑电实验室内进行,实验室隔音、隔光、安静、舒适。实验前给被试洗好头发并吹干,关闭通讯设备。让被试熟悉实验室环境,填写被试基本情况调查表。
实验在只有主试和被试的独立环境中进行,被试单独在房间里完成实验任务。首先向被试介绍有关实验的要求:实验过程中尽量放松全身,尤其是放松头部和面部肌肉,选舒服的坐姿坐好,实验正式开始后,头和身体尽量保持不动。将头放在面前的U型托上以保持头部不动,U型托距显示器的距离为60cm,双眼与显示器屏幕中心处于一条水平线上,双手舒服地放于小键盘的数字键上。告诉被试在实验开始后注意力尽量一直保持在前方显示屏上,并告知被试需要进行的简单操作及屏幕上呈现实验指导语。
实验指导语为:欢迎参加本实验。实验为两个部分,第一个部分为练习阶段。屏幕上会出现不同的发球视频,请你根据视频进行判断,并尽快按下相应的数字键进行反应,其中左手食指按“1”为转,右手食指按“3”为不转。练习共24次。练习结束后屏幕会播放短暂的发球视频,请根据发球动作判断来球的旋转方式,并尽快按相应的数字键进行反应。整个实验阶段请注意看屏幕,保持头部稳定。明白上述指导语后,请按任意键开始。
实验任务:首先出现1 000ms中央带“+”字符号的黑屏,然后播放发球录像视频,参加者根据视频进行判断,以最快的速度按相应键盘上的数字键:1,转;3,不转,视频呈现时间为2s,按键反应完成后,随后出现2s的黑屏,接着播放下一个随机视频文件,共160个视频,实验时间大约为20min(图1)。

图1 实验任务示意图Figure 1. Experimental Task
2.5 脑电的记录和数据的离线分析
脑电数据使用德国Brain Product公司生产的ERP记录系统进行记录(电极点 Ag/AgCl电极,带宽:0.01~100 Hz,采样率为:1 000Hz),采用按照国际标准10~20扩展系统64导电极帽。电极参考点的位置在AFz和Fz电极之间,接地的参考在Pz和Oz之间。右眼外侧安置电极记录水平眼电(HEOG),左眼下方安置电极记录垂直眼电(VEOG)。电极与头皮间阻抗保持在5kΩ以下,同时相同的带宽和采样率监控眼动。记录的单个序列脑电持续大约为5s。
对获得的脑电数据进行离线的预处理,主要包括以双耳乳突的平均电位为参考,去除眨眼伪迹,排除噪音干扰和其他伪迹,35Hz低通滤波,去除50Hz市电干扰,波幅大于±100μV视为伪迹自动剔除。对预处理后的脑电数据应用于MATALA 2009a平台进行相干性计算。
2.6 脑电相干性分析
脑电的相干性,是指任意给定频率两个信号之间的耦合程度[29]。从生理学的角度来说,脑电的相干性分析反映了大脑不同脑区之间的合作。相干性的值通过使用每个频率点λ来计算:

其中,fxy是两个信号x和y的互功率谱,fxx和fxy分别是信号x和y自功率谱。频域相干性是对两个信号在不同频率下的相位一致性的量度。事件相关相干性(event-related EEG coherence,ERCoh)的取值范围在0~1。当相干系数取1时,说明EEG信号y是x的在频段上的线性响应;相干系数为0时表明这两个EEG信号在某一频段上是不相干的,亦即两个信号是完全独立的。基于先前的研 究[4,5],具 体 的 两 个 电 极 点 之 间 的 相 关 通 过 MATLAB 2009a环境下编程实现。在相干函数计算时分段长度设置为1 024个样本点,重叠(overlap)设置为50%,短时傅里叶变换窗口类型(segment taper)设置为Hanning window。
基于前人的研究[5]和本研究的目的,对顶叶半球内(intra-hemisphere)的功能耦合进行评估,计算顶叶和其他脑区电极之间的脑电相干性,感兴趣的电极对为:P3-F3(左侧的顶叶和额叶),P3-C3(左侧的顶叶和中央脑区),P3-T7(左侧的顶叶和颞叶),P3-O1(左侧的顶叶和枕叶),P4-F4(右侧的顶叶-额区),P4-C4(右侧的顶叶和中央脑区),P4-T8(右侧的顶叶和颞叶),P4-O2(右侧的顶叶和枕叶脑区)。为了评估半球间(inter-hemisphere)的功能耦合,我们计算了 F3-F4,C3-C4,P3-P4,T7-T8,O1-O2电极之间的脑电相干性(图2)。脑电EEG功能耦合电极对的选取基于以下两点,第一,我们在10~20系统下选取的电极点计算的结果与其他前人研究中选取的那些电极点计算的结果可以进行比较[5,34];第二,所选电极对至少包括左右半球中感兴趣的视觉-空间脑区和躯体运动感觉区(左侧额区、右侧额区、左侧中央区、右侧中央区、左侧顶区、右侧顶区、左侧枕区、右侧枕区、左侧颞区、右侧颞区)。
2.7 数据的处理与分析
行为数据的采集采用E-prime 2.0软件包中E-Run运行程序并记录被试的反应时和反应正确率。E-DataAid对数据进行初步筛选和处理。E-Merge对不同组被试的数据进行合并。对异常数据进行筛选和剔除。行为数据的正确率,反应时间结果采用重复测量方差分析进行处理。
脑电数据的筛选和处理 实验中专家组和新手组各有两人在晚上进行的测试,由于晚上测试的两组被试脑电的波动较大,所以剔除了,最终处理的脑电数据专家组和新手组各为16人。根据本实验目的观察所有“转球”和“不转”正确识别的脑电数据进行处理,计算动作识别期间2s内的脑电相干性,即事件相关相干性(ERCoh)。主要统计目的是想探讨在动作识别过程中运动专家不同脑区功能耦合性。我们对不同频段(低频alpha、高频alpha、低频beta、高频beta)的脑电数据采用重复测量方差分析进行处理,在半球内脑内功能耦合的脑电方差分析以组别为组间变量,以旋转(转、不转)和脑区(左右顶-额、左右顶-中央区、左右顶-枕、左右顶-颞)为组内变量,进行2(组别)×2(转、不转)×8(脑区)的重复测量方差分析,感兴趣的电极对为 P3-F3、P4-F4、P3-C3、P4-C4、P3-O1、P4-O2、P3-T7和P4-T8。

图2 脑电电极位置图Figure 2. Electroencephalographic(EEG)Electrode Montage
半球间的功能耦合以组别为组间变量(专家、新手),以旋转(转、不转)和脑区(额区、中央区、顶区、枕区和颞区)为组内变量,进行2(组别)×2(旋转)×5(脑区)的重复测量方差分析,感兴趣的电极对有F3-F4(额区)、C3-C4(中央运动区)、P3-P4(顶区)、O1-O2(枕区)和T7-T8(颞区)。
所有重复测量方差分析使用Mauchley’s test的球型检验评估方差分析的齐性,对不满足球型检验的统计量采用Greenhouse-geisser法矫正自由度和P值,事后比较采用LSD法,α水平定义为P<0.05。按照Cohen标准[2],确定效应量η2p(effect size)达到0.15、0.06、0.01时,分别属于强效应、中等效应、弱效应。本研究对只有统计学意义的结果进行讨论分析。
3 结果
3.1 行为数据结果
对于行为数据以组别为组间变量,旋转为组内变量,分别以反应时间和反应正确率为因变量分别进行重复测量方差分析。首先对被试的反应时按照3个标准差法则进行了筛选,对反应时间和反应正确率数据进行KS检验,数据符合正态分布。不同水平运动员反应正确率的平均数和标准差见表1。
表1 本研究不同水平运动员发球动作识别正确率和反应时的平均数和标准差一览表Table 1 Means Value of Acc and RT for the Different Players during Action Judgment(±SD)

表1 本研究不同水平运动员发球动作识别正确率和反应时的平均数和标准差一览表Table 1 Means Value of Acc and RT for the Different Players during Action Judgment(±SD)
转不转X SD专家(n=16) 正确率(%)X SD 0.66 0.22 0.54 0.21反应时间(ms) 1 604.78 61.05 1 668.00 94.88新手(n=16) 正确率(%) 0.29 0.22 0.22 0.09反应时间(ms)1 621.93 152.53 1 613.24 198.09
对不同水平运动员发球动作识别的正确率方差分析结果显示,组别的主效应显著F(1,30)=33.942,P=0.000,=0.531,运 动 专 家 的 正 确 率 (X=0.600,SE=0.042)高于运动新手的正确率(X=0.256,SE=0.042);旋转的主效应 显著 F(1,30)=7.344,P=0.011,η2p=0.197,运动员识别转球的正确率(X=0.474,SE=0.039)大于识别不转球的正确率(X=0.381,SE=0.029);旋转和组别的交互作用不显著。反应时间的方差分析结果不存在任何显著性差异的主效应和交互作用。
以上结果表明,乒乓球运动员对于旋转动作的识别正确率存在水平差异,专家球员好于新手球员。两组运动员对于转球的识别好于不转球的识别,但在动作识别的反应时间上水平间不存在任何差异。
3.2 脑电相干性结果
3.2.1 专家和新手半球内不同频率相干性结果
低频alpha半球内重复测量方差分析结果显示,组间的主效应显著F(1,30)=23.030,P<0.01,η2p=0.434;旋转的主效应不显著 F(1,30)=0.009,P>0.05,η2p=0.015;脑区的主效应显著F(3.049,91.468)=80.624,P<0.01,η2p=0.729;脑区和组别的交互作用显著F(3.049,91.468)=2.918,P<0.05,η2p=0.089。交互作用后的简单效应检验结果显示在P3-C3(F=9.71,P=0.003)、P3-F3(F=17.11,P=0.000)、P3-O1(F=27.66,P=0.000)、P3-T7(F=30.47,P=0.000)、P4-F4(F=14.27,P=0.000)、P4-T8(F=10.19,P=0.002)电极对组别间存在显著性差异,乒乓球运动员的相干性显著低于乒乓球新手,且主要表现在左侧脑区。

表2 本研究半球内不同频段相干性方差分析一览表Table 2 ANOVA of the Intra-Hemispheric Different Frequency ERcoh

图3 本研究半球内不同频段运动专家(expert)和运动新手(novice)之间电极对脑电相干性交互作用示意图Figure 3. Mean Values of the Intra-Hemispheric Event-Related Coherence Amplitude in the Expert and Novices for the P3-C3,P4-C4,P3-F3,P4-F4,P3-O1,P4-O2,P3-T7,P4-T8,Electrode Pairs at Low-Frequency Alpha,High-Frequency Alpha,Low-Frequency Beta,High-Frequency Beta Bands
高频alpha半球内的重复测量方差结果显示,组别的主效应显著F(1,30)=15.670,P<0.01=0.343;旋转的 主 效 应 不 显 著F(1,30)=0.078,P>0.05,=0.003;脑区的主效应显著F(3.969,119.055)=101.425,P<0.01,=0.772;脑 区 和 组 别 的 交 互 作 用 显 著F(3.049,91.468)=4.111,P<0.05,=0.121。简单效应检验结果显示,P3-C3(F=10.53,P=0.002)、P3-F3(F=5.32,P=0.024)、P3-O1(F=21.47,P=0.000)、P3-T7(F=31.21,P=0.000)电极对组别间存在显著性差异,乒乓球运动员的相干性显著低于乒乓球新手。
低频beta半球内的重复测量方差结果显示,组间的主效应显著F(1,30)=6.355,P<0.05,=0.175,旋转的主效应不显著F(1,30)=0.267,P>0.05,=0.009,脑区的主效应显著F(3.044,119.055)=91.309,P<0.01,=0.734;脑 区 和 组 别 的 交 互 作 用 显 著F(3.044,119.055)=3.292,P<0.05=0.091。简单效应检验结果显示,P3-C3(F=7.57,P=0.008)、P3-F3(F=9.28,P=0.003)、P3-O1(F=7.59,P=0.008)、P3-T7(F=11.59,P=0.001)电极对组别间存在显著性差异,乒乓球运动员的相干性显著低于乒乓球新手。
高频beta半球内的重复测量方差结果显示,组间的主效应不显著F(1,30)=0.925,P>0.05=0.030,旋转的主效应不显著F(1,30)=0.628,P>0.05,=0.021,脑区的主效应显 著F(3.737,112.106)=70.935,P=0.000,0.703;脑区和组别的交互作用存在边缘性显著F(3.737,112.106)=2.371,P=0.061=0.073。交互作用后的简单效应检验结果显示,P3-F3(F=5.71,P=0.027)、P3-T7(F=5.32,P=0.024)电极对的组别间存在显著性差异,乒乓球运动员的相干性显著低于乒乓球新手。
以上半球内不同频段运动员发球动作识别的相干性结果显示,在低频alpha频段双侧的顶额叶(P3F3、P4F4)和顶颞叶(P3T7、P4T8)、左侧顶叶与左侧中央运动区(P3C3)、左侧顶叶与左侧枕叶(P3O1)电极对运动专家低于运动新手。高频alpha频段运动专家在左侧脑区的顶叶与左中央运动区(P3C3)、左额叶(P3F3)、左枕叶(P3O1)和左颞叶(P3T7)相干性低于运动新手;而在低频beta段内左侧顶叶与额叶、中央区和颞叶也具有相同的趋势,高频beta频段则只在左侧顶叶与左侧额叶(P3F3)、左侧颞叶(P3T7)脑区间存在差异。
3.2.2 专家和新手半球间不同频段的相干性结果
图4显示了专家和新手在发球动作识别时低频alpha、高频alpha、低频beta、高频beta不同频段内半球间F3-F4,C3-C4,P3-P4,O1-O2,T7-T8电极对的脑电相干性,从图中可以看出,运动专家动作识别时半球间不同脑区电极对相干性低于运动新手。表3显示了半球间不同频段组别与旋转、脑区电极对的相干性重复测量方差分析结果。
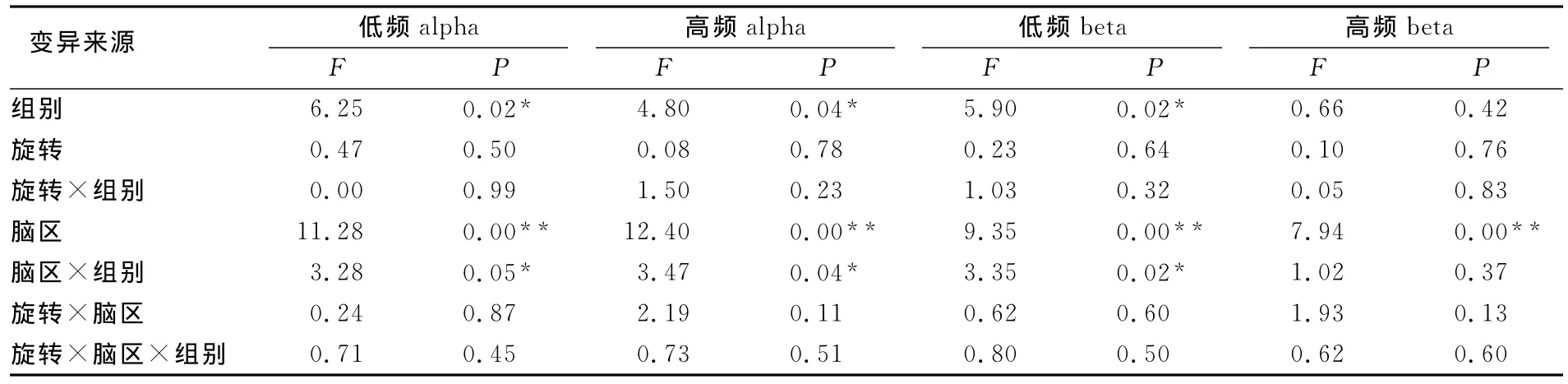
表3 本研究半球间不同频段相干性方差分析一览表Table 3 ANOVA of the Inter-Hemispheric Different Frequency ERcoh
低频alpha半球间不同脑区电极对的重复测量方差结果显示,组别的主效应显著F(1,30)=6.246,P<0.05,=0.172;旋转的主效应不显著F(1,30)=0.468,P>0.05,=0.015,脑区的主效应显著F(2.765,82.958)=11.280,P<0.01=0.273。脑区和组别之间存在交互作用F(2.765,82.958)=3.280,P<0.05,=0.113。简单效应检验发现在C3-C4(F=8.33,P=0.005),F3-F4(F=4.28,P=0.043),O1-O2(F=4.29,P=0.043),P3-P4(F=6.85,P=0.011)电极对组别之间的相干性存在显著性差异。
全国青少年校园足球工作始于2009年。自2014年11月26日国务院召开全国青少年校园足球工作电视电话会议后,教育部正式牵头负责全国青少年校园足球工作。3年多来,教育部一件事情接着一件事情干,使得校园足球工作成绩斐然,也为学校体育工作指出了新方向、开辟了新思路、创建了新模式。
高频alpha半球间不同脑区电极对的重复测量方差结果显示,组别的主效应显著F(1,30)=4.803,P<0.05,=0.138,运动专家的相干性(0.501±0.008)显著低于运动新手的相干性(0.527±0.008)。旋转的主效应不显著F(1,30)=0.078,P>0.05=0.003,脑区的主效应显著F(3.037,91.105)=12.470,P<0.01=0.294。脑区与组别的交互作用显著F(3.037,91.105)=3.470,P<0.05,=0.094。 简 单 效 应 检 验 发 现,在 C3-C4(F=6.95,P=0.002),P3-P4(F=3.40,P=0.045)电极对组别之间的相干性差异显著,乒乓球运动员的相干性显著低于运动新手。
低频beta半球间不同脑区电极对的重复测量方差结果显示,组别的主效应显著F(1,30)=5.898,P<0.05,=0.164,运动专家的相干性(0.470±0.011)显著低于运动新手的相干性(0.506±0.011)。旋转的主效应不显著F(1,30)=0.228,P>0.05=0.008,脑区的主效应显著F(2.146,64.380)=9.351,P<0.01,=0.238,脑区与组别的交互作用显著F(2.146,64.380)=3.351,P<0.05,=0.108,简单效应检验结果显示,在 C3-C4(F=7.95,P=0.006)P3-P4(F=6.48,P=0.013)电极对组别之间的相干性存在显著性差异,乒乓球运动员的相干性显著低于运动新手。
高频beta半球间不同脑区电极对的重复测量方差结果显示,组别的主效应不显著F(1,30)=0.657,P>0.05,=0.021,旋转的主效应不显著F(1,30)=0.095,P>0.05,=0.003,脑区的主效应显著F(2.318,69.544)=7.943,P<0.01,=0.209。脑区与旋转和水平之间不存在任何的交互作用。脑区主效应多重比较结果显示,颞叶与运动区、额叶和顶叶均存在显著性异(P<0.01),其中颞叶(T7T8)的相干性最低,额叶(F3F4)的相干性最高。
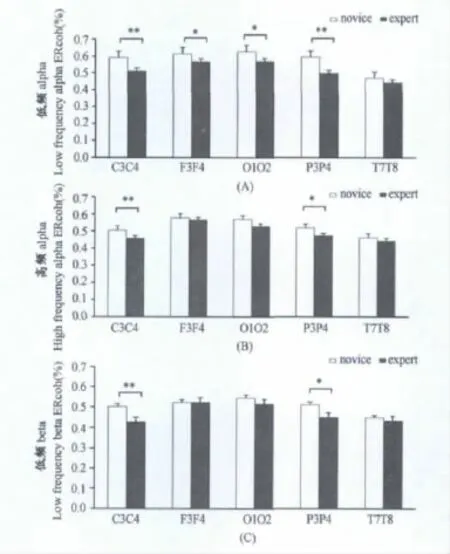
图4 本研究半球内不同频段不同脑区电极对运动专家(expert)和运动新手(novice)之间的脑电相干性交互作用示意图Figure 4. Mean Values of the Inter-Hemispheric Event-Related Coherence Amplitude in the Expert and Novices for the F3-F4,C3-C4,P3-P4,O1-O2,T7-T8,Electrode Pairs at Low-Frequency Alpha,High-Frequency Alpha,Low-Frequency Beta,High-Frequency Beta Bands
半球间脑区的相干性结果显示,低频alpha频段在顶叶、额叶、枕叶和中央区,运动专家与运动新手的相干性差异性显著,运动专家低于运动新手,高频alpha和低频beta频段在中央运动区(C3C4)和顶叶(P3P4)存在差异,高频beta频段在组间差异不显著。
4 讨论
人们希望从不同脑区电极对的记录信号中发现同频率固定相位的振荡,由此揭示不同脑区之间在一定认知状态下的协作方式[1]。本研究采用脑电的相干性分析技术,探讨乒乓球运动员进行发球动作识别时皮层与皮层之间神经元同步振荡的功能耦合性,以评估乒乓球运动员动作识别时的神经效率和大脑皮层的功能耦合特征。
4.1 乒乓球运动员动作识别时神经效率
运动专家的完美操作与高相干性相联系,还是表现为低相干性呢?有研究认为,高相干性反映了两个不同脑区之间的连接,而低相干性则反映了脑区间的自动化和精细化(Refinement)[33]。本研究结果显示,与乒乓球新手相比,乒乓球运动员在较好的操作中表现为脑区间的低相干性。Deeny等(2003,2009)对射击运动员和射击新手研究表明,专家与新手相比展现了低相干性,特别是左侧颞叶与中央运动区,显示是运动专家执行专项任务时,减少了参与运动过程的认知加工,支持运动专家皮层网络的精细分化,表明了专家与新手在运动计划相关的记忆过程和视觉空间线索执行上的不同。在视觉空间、语言和运动皮层之间低相干性代表了运动员专业化的增加和运动技能变得更加精细[13]。事实上在运动技能学习和发展研究中也显示,随着运动技能的学习,脑区之间的相干性降低[13]。低水平阶段运动新手皮层活动特征是在认知任务中相关和不相关的脑区都产生激活[19],反映了相对的不稳定的神经过程,然而随着技能水平的提高,动作变得更加精细、稳定和自动化,相关任务脑区不必要功能连接降低。Oliviero等(2003)使用TMS对手部运动训练后引起的皮层之间的相干性分析显示,训练后在高频alpha频段半球内中央运动区的相干性降低[28]。本研究结果表明与运动新手相比,乒乓球运动员发球动作识别时大脑相关皮层之间的低相干性,反映了乒乓球运动员长期的运动技能学习使他们大脑皮层之间功能更加自动化和精细分化,相关脑区较低的神经元同步化振荡就可以完成发球旋转的识别,体现了神经高效性。
4.2 乒乓球运动员动作识别时脑区功能组织
本研究结果显示,与乒乓球新手相比,乒乓球运动员动作识别时半球内低频alpha双侧顶叶与额叶、颞叶,左侧顶叶与左侧枕叶展现了低相干性,高频alpha在左侧顶叶与额叶、枕叶和颞叶展现了低相干性,而半球间乒乓球运动员低频alpha在中央运动区、额叶、顶叶、枕叶和颞叶的相干性低,高频alpha在顶叶和中央运动区也展现了低相干性,即在alpha频段展现了大脑皮层间广泛的功能耦合。目前的研究结果对运动员大脑的功能组织具有怎样的生理意义呢?Neubauer(1995)认为,alpha(8~13Hz)和低频beta(13~22Hz)反映了总体的皮层与皮层之间的连接,而高频(如36~44Hz)则代表了具体脑区的加工过程,或与具体任务的脑区激活有关[27]。也有研究认为,alpha频率反映了基底前脑、背侧丘脑和皮层环路对感觉运动区和大脑认知信息传递和检索的促进或抑制[7],且低频alpha和高频alpha也具有不同的意义,低频alpha被认为与整体大脑唤醒或研究对象的注意状态和努力有关,而高频alpha反映了与具体任务相关的躯体或语义信息的神经系统管理[22]。乒乓球运动员发球动作识别时alpha频段脑区间广泛的低相干性,可能说明,乒乓球运动员只需要较低的唤醒水平和注意努力,就可以完成发球动作识别信息的检索和传递,其调节的方式是通过低频alpha调节大脑整体的注意状态,通过高频alpha调节动作识别任务的“视觉-空间”信息和任务执行的过程。涉及的主要脑区为负责“视觉-空间”的后顶叶与额叶皮层(P3-F3,P4-F4)、颞叶皮层(P3-T7,P4-T8)和中央运动区(C3-C4)。
4.3 乒乓球运动员动作识别脑区功能耦合
乒乓球运动员发球动作识别的过程是基于感觉信息与长时记忆中的信息进行比较,并决定与记忆中的模式进行最佳匹配的过程,该过程不仅依赖于视觉信息的获取,也依赖于运动员具有的专项知识和经验。研究结果显示,乒乓球运动员正确识别时alpha频率双侧额顶叶、颞顶叶的功能耦合与Del Percio等研究相一致,该研究认为,较好的运动操作与双侧alpha的调节有关。前人的研究认为,后顶叶皮层和额叶皮层对控制视觉空间注意自上而下的加工起着关键的作用[31]。空间注意的生理机制是视觉皮层神经元群之间的同步振荡活动[17],这也是alpha节律的主要调节机制,反映了大脑皮层椎体神经元围绕着的皮层之间和丘脑动态的同步整合活动[29]。乒乓球运动员在动作识别过程中顶叶-额叶皮层的功能耦合反映了运动员在动作识别过程中对发球信息的空间注意显示了明显的专长优势。认知加工过程中顶叶与颞叶的功能耦合是通过调节颞叶提取的与任务相关的特征信息进行整合实现的[12],乒乓球运动员强大的记忆表征已经建立好一个很好的内部模式,因此,在发球动作识别过程中顶叶和颞叶之间显示了更低的相干性。乒乓球运动员基于顶叶以这种网络的方式建立了关于发球动作识别任务信息的整体感知。在动作识别过程中乒乓球运动员依赖于已建立好的内部模式来调节信息的匹配和执行,启动了自上而下的加工。而乒乓球新手额顶叶和颞顶叶的高相干性可能与新手运动员对于发球旋转的识别任务,没有形成内部的记忆表征,更多的依赖于环境中的信息线索,启动了自下而上的加工,而不是启动记忆来面对当前的任务,这也说明了顶叶-额叶和顶叶-颞叶皮层耦合在动作识别的匹配过程起着关键的作用。
此外,研究结果显示与乒乓球新手相比,乒乓球运动员在alpha频段和低频beta频段内左侧顶叶与左侧枕叶、左侧中央运动区也显示了功能的耦合,在低频beta频段内中央运动区和顶叶表现出低相干性。研究者认为,顶枕叶脑区alpha频段在静息状态时同步化振荡增加,而在视觉目标的预期和编码过程中同步化振荡降低[32],因此,乒乓球运动员发球动作识别时顶枕区的低相干性可能暗示了与运动新手相比,乒乓球运动员视觉目标的注意和编码的改善。中央运动区调节运动皮层的活动,而运动活动又受运动计划脑区(FZ)和运动执行脑区(CZ)的共同调节,顶叶和中央运动区功能耦合反映了与整体注意有关神经元的募集。有研究显示,顶叶与枕叶、中央区的相干性与更好视觉运动操作绩效相联系[4]。在运动保持任务中beta频率会在初级运动皮质、运动皮质和顶叶进行神经元同步振荡[6]。在视觉运动任务中beta(15~35Hz)频率的相干性反映了感觉运动皮层的有效连贯性[26]。以视听刺激为材料的研究中也表明,beta频段的同步性对视觉和运动脑区的有效功能整合起着重要作用[21]。基于以上研究,本研究中乒乓球运动员beta频段的低相干性,暗示了乒乓球运动员动作识别时运动皮层和顶叶皮层之间已经建立了高效连接,故神经元同步化振荡幅值较低。乒乓球运动员在发球动作识别过程中这些脑区的功能耦合,也说明了发球动作的识别是一个复杂的认知过程,涉及多个脑区的分工与合作。
研究结果显示,半球内不同频段内均显示了左侧半球相关脑区的功能耦合。有研究认为,在高水平的运动操作中左半球认知过程起着关键作用[22]。大脑半球偏侧化研究显示,精细的运动控制和语言倾向于定位左半球(对右利手而言),空间定位能力倾向于定位右半球。对“裂脑”人的研究证实左半球倾向于模式识别,而右半球更倾向于进行空间分析[25]。本研究的实验对象均为右利手运动员,并进行发球动作的识别任务,所以,本研究也证实了大脑左侧半球在乒乓球运动员发球动作识别过程中起着重要作用。
5 结论
1.乒乓球运动员发球动作识别时脑区间的相干性低,脑区间分工更加精细化,体现了神经效率。
2.Alpha频段神经元的同步化振荡在乒乓球运动员动作识别时起着重要的调节作用。
3.乒乓球运动员发球动作识别时在额顶叶、颞顶叶、枕顶叶和中央运动区显示了功能耦合,并表现为左侧半球优势。
4.乒乓球运动员在动作识别的编码和匹配识别阶段存在认知优势。
[1]洪波,杨福生,岳小敏,等.基于多变量AR模型的脑电相干性分析及其在脑区协作机制研究中的应用[J].生物物理学报,2001,17(1):105-113.
[2]全朝鲁.效果量的意义及测定方法[J].心理学探新,2003,86(2):39-44.
[3]ALBRECHT J RILK,SURJIO R SOEKADAR,PAUL SAUSENG,et al.Alpha coherence predicts accuracy during a visuomotor tracking task[J].Neuropsychol,2011,49(13):3704-3709.
[4]BABILONI C,BRANCUCCI A,VECCHIO F,et al.Anticipation of somatosensory and motor events increases centro-parietal functional coupling:an EEG coherence study[J].Clin Neurophysiol,2006,117(5):1000-1008.
[5]BABILONI C,VECCHIO F,CAPPA S,et al.Functional frontoparietal connectivity during encoding and retrieval processes follows HERA model.A high resolution study[J].Brain Res Bulletin,2006b,68(4):203-212.
[6]BROVELLI A,DING M,LEDBERG A,et al.Beta oscillations in a large-scale sensorimotor cortical network:directional influences revealed by Granger causality[J].Proceedings National Academy Sci U S A,2004,101(26):9849-9854.
[7]BRUNIA C.Neural aspects of anticipatory behavior[J].Acta Psychol,1999,101(2-3):213-242.
[8]BUSK J,GALBRAITH G C.EEG correlates of visual-motor practice in man[J].Electroencephalography Clin Neruo Physiol,1975,38(4):415-422.
[9]CLAUDIO BABILONI,FRANCESCO INFARINATO,NICOLA MARZANO.Intra-hemispheric functional coupling of alpha rhythms is related to golfer’s performance:A coherence EEG study[J].Int J Psychophy,2011,82(3):260-268.
[10]C GERLOFF,J RICHARD,J HADLEY,et al.Functional coupling and regional activation of human cortical motor areas during simple,internally paced and externally paced finger movements[J].Brain,1998,121(8):1513-1531.
[11]CICEK M,NALCACI E.Inter-hemispheric asymmetry of EEG alpha activity at rest and during the Wisconsin card sorting test:relations with performance[J].Biological Psychol,2001,58(1):75-88.
[12]DEENY S P,HILLMAN C H,JANELLE,et al.Cortico-cortical communication and superior performance in skilled marksmen:An EEG coherence analysis[J].J Sport Exe Psychol,2003,25(2):188-204.
[13]DEENY S P,AMY J HAUFLER,MARK SAFFER,et al.E-lectroencephalographic coherence during visuomotor performance:A comparison of cortico-cortical communication in experts and novices[J].J Motor Behavior,2009,41(2):106-116.
[14]DEL PERCIO C,IACOBONI M,LIZIO R,MARZANO N,et al.Functional coupling of parietal alpha rhythms is enhanced in athletes before visuomotor performance:A coherence electroencephalographic study[J].Neurosci,2011,175:198-211.
[15]DEL PERCIO C,ROSSINI PM,MARZANO N,et al.Is there a“neural efficiency”in athletes?A high-resolution EEG study[J].Neuroimage,2008,42(4):1544-1553.
[16]DEL PERCIO C,BABILONI C,MARZANO N,et al.“Neural efficiency”of athletes’brain for upright standing:a high-resolution EEG study[J].Brain Res Bull,2009b,79(3-4):193-200.
[17]FRIES P.A mechanism for cognitive dynamics:neuronal communication through neuronal coherence[J].Trends Cognitive Sci,2005,9(10):474-480.
[18]GEVINS A,SMITH ME,LEONG H,MCEVOY L,et al.Monitoring working memory load during computer-based tasks with EEG pattern recognition methods[J].J Human Factors Ergonomics Soc,1998,40(1):79-91.
[19]HATFIELD B D,HILLMAN C H.The psychophysiology of sport:A mechanistic understanding of the psychology of superior performance[M].In R Singer,H Hausenblas,C Janelle(Eds),Handbook Sport Psychol,New York:Wiley Sons,2001.
[20]HAUFLER A J,SPALDING T W,SANTA MARIA,et al.Erratum to“Neuro-cognitive activity during a selfpaced visuospatial task:Comparative EEG profiles in marksmen and novice shooters.”[J].Biological Psychol,2002,59(1):87-88.
[21]HIPP J F,ENGEL A K,SIEGEL M.Oscillatory synchronization in large-scale cortical networks predicts perception [J].Neuron,2011,69(2):387-396.
[22]KERICJ S E,MCDOWELL K,HUNG T,et al.The role of the left temporal region under the cognitive motor demands of shooting in skilled marksmen [J].Biological Psychol,2001,58(3):263-277.
[23]KLIMESCH W.EEG alpha and theta oscillations reflect cognitive and memory performance:a review and analysis[J].Brain Res Rev,1999,29(2-3):169-195.
[24]LOVATELLI T,CURSI M,LIBERATI D,et al.EEG coherence in Alzheimer’s disease[J].Electroencephalography Clin Neurophysiol,1998,106(3):229-237.
[25]MARGARET G FUNNELL,PAUL M CORBALLIS,MICHAELl S GAZZANIGA.A depcit in perceptual matching in the left hemisphere of a callosotomy patient[J].Neuropsychol,1999,37(10):1143-1154.
[26]MATTHIAS WITTE,LUIS PATINOL,AGNIESZKA ANDRYKIEWICZ.Modulation of human corticomuscular betarange coherence with low-level static forces[J].Eur J Neuroscience,2007,26(12):3564-3570.
[27]NEUBAUER A C,FREUDENTHALER H H,PFURTSCHELLER G.Intelligence and spatio-temporal patterns of event-related desynchronization[J].Intelligence,1995,20(3):249-267.
[28]OLIVIERO A,STRENS L H,DI LAZZARO V,et al.Persis-tent effects of high frequency repetitive TMS on the coupling between motor areas in the human[J].Experimental Brain Res,2003,149(1):107-113.
[29]PFURTSCHELLER G,LOPES DA SILVA F H.Event-related EEG/MEG synchronization and resynchronization:basic principles[J].Clin Neurophysiol,1999,110(11):1842-1857.
[30]RAPPELSBERGER P,PETSCHE H .Probability mapping:power and coherence analyses of cognitive processes[J].Brain Topography,1988,1(1):46-54.
[31]SERENCES J T,YANTIS S.Selective visual attention and perceptual coherence[J].Trends Cognitive Sci,2006,10(1):38-45.
[32]SAUSENG P,KLIMESCH W,STADLER W,et al.A shift of visual spatial attention is selectively associated with human EEG alpha activity[J].Eur J Neuroscience,2005,22(11):2917-2926.
[33]WEISS S,MUEELLER H M.The contribution of EEG coherence to the investigation of language[J].Brain and Language,2003,85(2):325-343.
[34]ZHU F F,MAXWELL J P,HU Y,et al.EEG activity during the verbal-cognitive stage of motor skill acquisition[J].Biological Psychol,2010,84(2):221-227.
