新疆野扁桃与栽培扁桃花原基解剖结构观察
2013-04-10钟海霞刘立强
钟海霞 ,陆 婷 ,刘立强 ,李 疆 ,张 磊 ,何 龙
(新疆农业大学a.林学与园艺学院;b.科研处,新疆 乌鲁木齐 830052)
扁桃A. communisL.又名普通扁桃、栽培扁桃、巴旦杏,是蔷薇科Roseceae桃属Amygdalus植物[l]。扁桃是落叶乔木,萌芽力强,嫁接苗在第3年开始结果,6~8年后可大量结果,结果寿命长达100 a以上,经济效益显著[2]。扁桃品种具有较强的适应性,栽植密度、修剪技术、花期管理、病虫害防治等因素对扁桃的高产优质栽培具有巨大的影响[3]。我国的扁桃现在有6个种,野扁桃Amygdalus ledebourianaSchleche又称野巴旦,是扁桃的一个种。
新疆是世界32个扁桃产区之一,是我国扁桃种植规模最大的产区。扁桃花芽在休眠期易受冻害,如2008年冬季莎车县的低温冻害使当地扁桃产量减产70 %以上[4],严重影响了当地经济的发展。因此,选育抗寒性强的扁桃品种是促进扁桃产业化发展的必由之路,利用野生抗寒种质资源对改良栽培品种具有重要意义。新疆塔城地区裕民县约有400 hm2的野扁桃林自然保护区,是目前世界上最大的野生扁桃生长区[5],该地区1月平均气温为-10 ℃左右,绝对最低温度为-34.5℃[6],而野扁桃A. ledebourianaschleche仍能安全越冬。目前已有许多学者对扁桃进行了研究,李疆[7]对扁桃的结实率低的生理与分子机制进行了研究,结果表明:其制约因素有扁桃的树体营养和激素、花芽分化、坐果和果实发育的生物学特性、S基因型等;徐叶挺等人[8]研究发现,新疆野扁桃花芽形态分化于7月上旬~9月上旬,分为5个时期,即花芽分化初期、花萼分化期、花瓣分化期、雄蕊分化期和雌蕊分化期;杨晓宇[9]和李斌等人[10]从生理生化方面进行研究,结果发现,野扁桃的抗寒性明显强于栽培扁桃;郭长奎等人[11]在研究扁桃自交不亲和的机制时获得了一个扁桃花粉的SFB基因,即花粉S决定子;王森等人[12-14]研究发现,不同盐分环境对扁桃凝胶弹性、粘附性、回复性等方面有一定的影响,还得出了美国扁桃花被细胞膜系统在-6 ℃时会崩溃,从而会伤害到雄蕊和雌蕊原基的结论,并将7个美国品种引种至河南南阳,观测其植物学及生长结果特性。一些学者研究发现,植物形态解剖结构与抗寒性存在密切关系且受环境因素影响较小[15-18]。鉴于此,文中对越冬前野扁桃和3个栽培扁桃品种的花原基进行了解剖学研究,试图通过分析比较野扁桃与栽培扁桃花原基的结构特征,探讨其与抗寒性的关系,以期为新疆扁桃高产优质栽培及抗寒种质资源的选育提供理论依据。
1 材料与方法
1.1 材 料
供试材料有野扁桃和栽培扁桃。野扁桃于2010年11月底采自新疆裕民县巴尔鲁克山野扁桃自然保护区;栽培扁桃有3个品种,从美国引进的栽培品种浓帕烈(Nonpareil)采自莎车县阿热斯兰巴格乡,新疆喀什地区的纸皮和矮丰这2个主栽品种则采自莎车县阿热勒乡。树龄为8~20 a,树势中等。
1.2 方 法
1.2.1 试验方法
从采集的各个品种的试验样品中选择饱满度好的花芽60~80个,剥去外部鳞片后用FAA固定液(70%酒精∶甲醛∶冰醋酸=89∶6∶5)固定,爱氏苏木精整染,用常规石蜡切片法制片[8],切片厚度8~10 μm,加拿大树胶封片,Motic Type 102M显微成像系统观察并拍照。Motic Images Advanced 3.2软件测量,切片数据取30个测量值的平均值。
1.2.2 数据统计分析
试验数据采用Excel和SPSS 17.0统计软件进行分析,采用Duncan’s新复极差法进行不同指标及品种之间的多重比较分析。
2 结果与分析
2.1 花萼原基解剖结构的观察与比较
对野扁桃与3个栽培扁桃品种的花萼原基进行形态解剖,用Motic Type 102M显微镜观察并拍照,每个品种选30张品质高的照片,应用Motic Images Advanced 3.2软件测量花萼厚度、细胞纵径、细胞横径、细胞间隙,结果如表1所示。表1表明,野扁桃与栽培扁桃花萼原基的细胞形态存在较大差别。野扁桃花萼原基细胞形状呈长椭圆形,排列紧密,栽培扁桃花萼原基细胞呈近圆形或方形,细胞排列不及野扁桃整齐(见图1)。野扁桃花萼原基细胞纵径和横径均极显著小于3个栽培扁桃细胞,野扁桃花萼原基厚度和细胞间隙极显著小于栽培品种(见表1)。

表1 野扁桃与栽培扁桃花萼原基解剖结构的观测结果†Table 1 Observation result of anatomical structures of the sepal primordia in A. Ledebouriana and A. communis µm
2.2 花瓣原基解剖结构的观察与比较
对野扁桃与栽培扁桃的花瓣原基进行形态解剖,用显微系统拍照,对花瓣厚度、花瓣上表皮、中层、下表皮的细胞纵径、细胞横径、细胞间隙进行测量,结果如表2所示。表2表明,野扁桃与栽培扁桃花瓣原基上表皮、中层、下表皮细胞形态有明显的不同。野扁桃上、下表皮细胞为矩长方形,栽培扁桃多为近方形;野扁桃中层细胞呈近方形,栽培扁桃为矩圆形(见图2)。由表2可知,野扁桃花瓣厚度均极显著小于3个栽培品种。野扁桃花瓣原基上表皮、中层、下表皮细胞的纵径、横径均极显著小于栽培品种,野扁桃花瓣上、中、下表皮细胞间隙与栽培品种的差异达到极显著水平,即均极显著小于3个栽培品种。
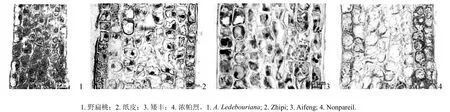
图1 野扁桃与栽培扁桃的花萼原基纵切面解剖图(100×)Fig.1 Vertical section anatomies of the sepal primordia in A. Ledebouriana and A. communis (100×)
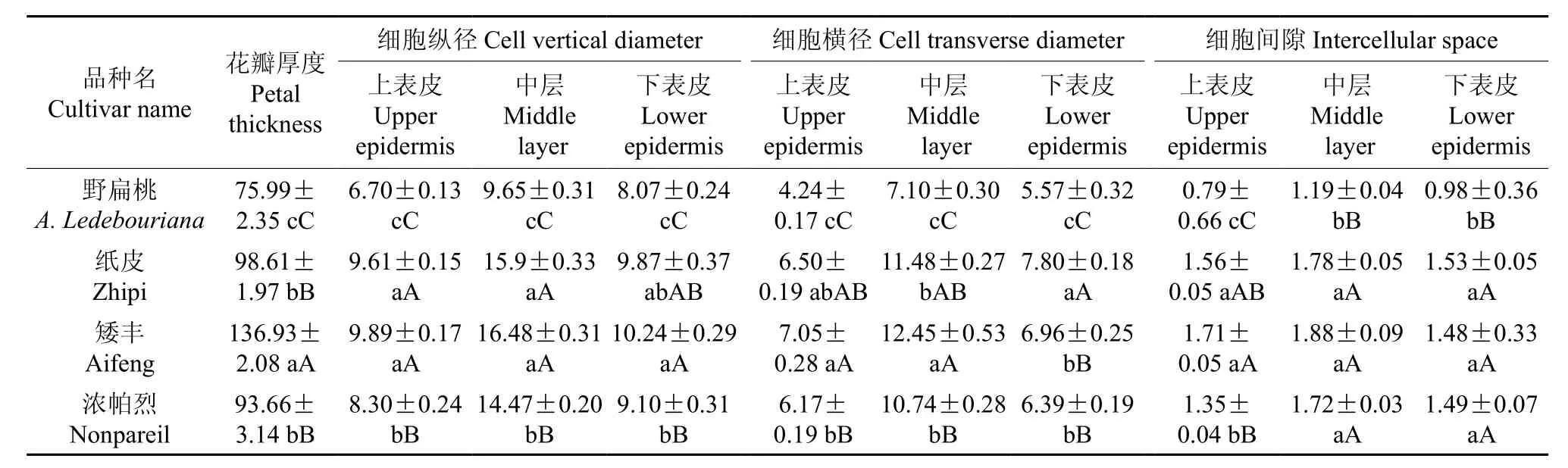
表2 野扁桃与栽培扁桃花瓣原基解剖结构的观测结果Table 2 Observation result of anatomical structures of the petal primordia in A. Ledebouriana and A. communis µm
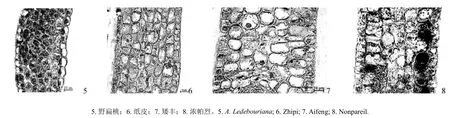
图2 野扁桃与栽培扁桃的花瓣原基纵切面解剖图(100×)Fig.2 Vertical section anatomies of the petal primordia in A. Ledebouriana and A. communis (100×)
2.3 花药原基解剖结构的观察与比较
对野扁桃与栽培扁桃的花药原基进行形态解剖并观察,应用软件测量花药的细胞纵径、细胞横径、细胞间隙、药壁厚度,结果如表3所示。表3表明,野扁桃与栽培扁桃花药原基的细胞形态和发育进程有着明显的区别。野扁桃和栽培扁桃花药的药壁为3~4层细胞组成,环状整齐排列;花药绒毡层细胞为多核细胞,细胞质浓,其发育为同时形。同一视野下,野扁桃花药原基与栽培品种的解剖结构有所不同,其发育进程比栽培扁桃慢,即野扁桃花原基越冬之前处于发育小孢子母细胞或更早期,但栽培品种已发育至四分体甚至达单核花粉期(见图3)。野扁桃花药原基的药壁细胞呈长方形,细胞核染色深,而栽培扁桃的药壁细胞则呈不规则长方形,细胞排列不及野扁桃规则。野扁桃花药原基中药壁的细胞纵径极显著小于栽培品种,细胞横径显著小于引进栽培品种,极显著小于当地主栽品种;野扁桃药壁的厚度和细胞间隙均极显著小于3个栽培品种(见表3)。
2.4 野扁桃与栽培扁桃雌蕊原基解剖结构的观察与比较
对野扁桃与纸皮、矮丰、浓帕烈的雌蕊原基花柱部分制作石蜡切片,Motic Type 102M显微镜观察并拍照,应用测量软件对花柱表皮和薄壁组织的细胞纵径、细胞横径、细胞间隙、花柱厚度进行测定,结果如表4所示。表4表明,野扁桃和栽培扁桃雌蕊原基花柱部分由表皮、薄壁组织和花柱通道组成,其细胞形态有所不同。野扁桃花柱表皮细胞呈长方形,薄壁组织细胞为规则多边形,细胞排列整齐,栽培扁桃花柱的表皮细胞呈长柱形,薄壁组织细胞呈不规则多边形,且不及野扁桃的花柱细胞排列紧密(见图4)。由表4可知,野扁桃花柱表皮和薄壁组织的细胞纵径和横径均极显著小于栽培品种,野扁桃花柱厚度和细胞间隙与栽培品种间的差异达到了极显著水平,即极显著小于栽培扁桃。

表3 野扁桃与栽培扁桃花药原基解剖结构的观测结果Table 3 Observation result of anatomical structures of the anther primordia in A. Ledebouriana and A. communis µm
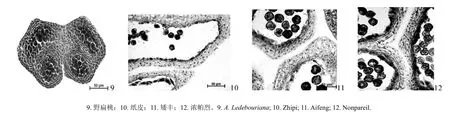
图3 野扁桃与栽培扁桃的花药原基纵切面解剖图(40×)Fig.3 Vertical section anatomies of the anther primordia in A. Ledebouriana and A. communis (40×)
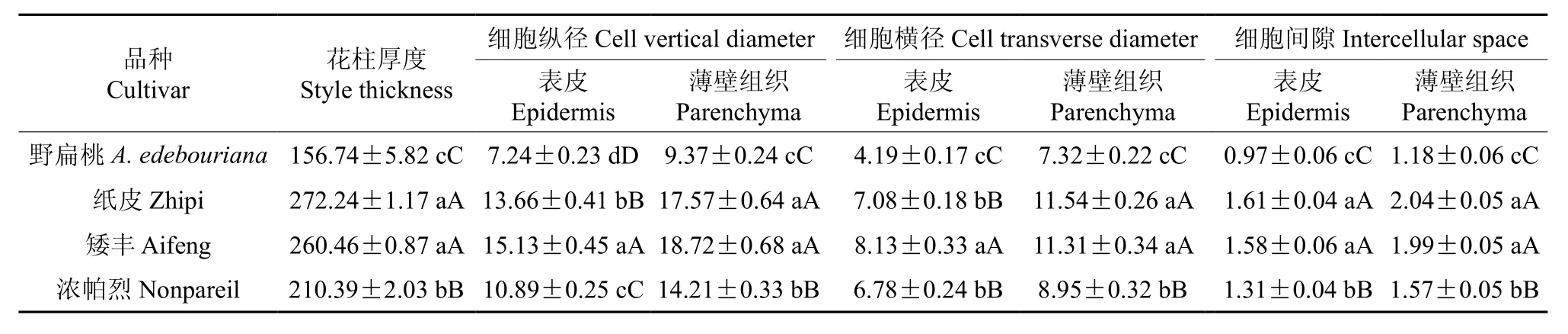
表4 野扁桃与栽培扁桃雌蕊原基解剖结构的观测结果Table 4 Observation result of anatomical structures of the pistil primordia in A. Ledebouriana and A. communis µm
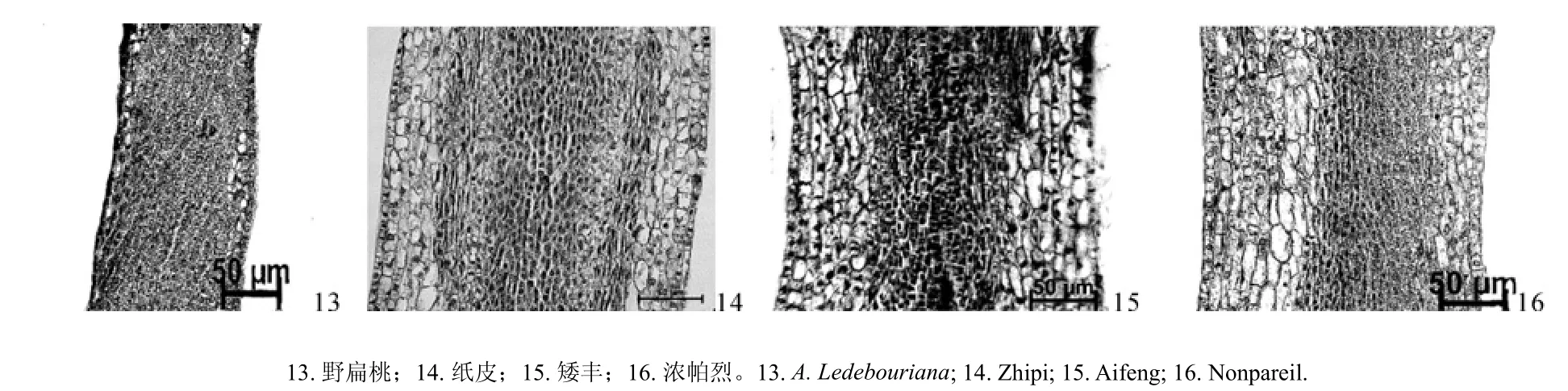
图4 野扁桃与栽培扁桃的雌蕊原基(花柱部分)纵切面解剖图(40×)Fig.4 Vertical section anatomies of the pistil primordia (style) in A. Ledebouriana and A. communis (40×)
3 结论与讨论
大量研究结果表明,植物的抗寒性与解剖结构有着密切关系,抗寒性强的品种其细胞较小、细胞排列紧密度大、疏松度小[19-20]、细胞间隙较小[21]、细胞出现严重的质壁分离现象[22],细胞核和线粒体嵴消失快[23-24],用其研究植物的组织结构可反映出植物对特定环境的适应性。
Herminda Reinoso等人[25]研究发现,果树的花器官原基细胞形态在休眠期发生了一系列变化,因而将相应花原基组织构造特征作为衡量扁桃抗寒性强弱的机理之一是比较客观的,自然越冬前野扁桃和栽培扁桃不同品种的受害症状最能反映出其自身的遗传本质。文中试验结果表明,抗寒性强的野扁桃花原基细胞纵径、横径均小,细胞排列紧密,细胞间隙小;而抗寒性弱的栽培扁桃花原基细胞纵径、横径较大,细胞排列较疏松,细胞间隙较大,这与房用等人[26]对茶树抗寒性研究得出的抗寒性强的茶树品种叶片的栅栏组织细胞较小的研究结果一致;与唐立红等人[27]研究得出的叶片组织紧密度大、疏松度小的品种抗寒性较强的研究结果一致;与郑家基等人[18]研究得出的细胞间隙小、利于提高植物抗寒能力的这一论断也相一致。此外,文中试验发现,抗寒性强的野扁桃花原基厚度小,而抗寒性弱的栽培品种花原基厚度大,这与邹雪娟等人[28]研究得出的结果不同,可能与所研究的材料及取材时期以及鳞片的结构和厚度不同有关。对于扁桃花原基厚度与其抗寒性的关系,在本试验中未显示出一定的变化规律,这一点还有待于进一步的研究。
文中初步探讨了扁桃花原基解剖结构与其抗寒性的关系,试验结果为扁桃抗寒性的鉴定提供了一定的参考依据。在生产实践中,可用新疆野扁桃作砧木进行嫁接,以提高栽培扁桃的抗寒性,扩大扁桃的栽培区域,进一步提高其经济效益。
参考文献:
[1]CALILBRNI A I. University of California Almond production manual [M]. California: Division of Agriculture and Natural Resources Publication,1996:3-24.
[2]李 疆,胡芳名,张智俊,等.扁桃主要生物学特性的观测[J].经济林研究,2003,21(3):39-40.
[3]张文越,史作安,张玉贞,等.扁桃引种及良种筛选试验[J].经济林研究,2011,29(1):76-84.
[4]杨 波,龚 鹏,车玉红. 6个美国扁桃品种在新疆喀什地区的引种适应性[J].西北农业学报,2012,21(1):115-120.
[5]李 疆,曾 斌,罗淑萍,等.我国野扁桃资源的保护及引种繁育[J].新疆农业科学,2006,43(1):61-62.
[6]史 征,宏 伟,郭 荔.野巴旦杏树造林技术[J].林业科技开发,2004,18(4):65.
[7]李 疆.新疆扁桃结实率低的生理与分子机制[J].经济林研究,2011,29(2):1-7.
[8]徐叶挺,李 疆,罗淑萍,等.野生巴旦杏花芽形态的分化过程[J].经济林研究,2008,26(4):86-89.
[9]杨晓宇.扁桃枝叶生理指标与抗寒性相关性研究[D].太原:山西大学, 2010.
[10]李 斌,刘立强,罗淑萍,等.扁桃花芽的抗寒性测定与综合评价[J].经济林研究,2012,30(3):16-21.
[11]郭长奎,李 疆,罗淑萍,等.扁桃花粉SFB基因的鉴定和序列分析[J].经济林研究,2009,27(3):18-23.
[12]王 森,谢碧霞,钟秋平,等.不同盐分环境对扁桃凝胶质构特性的影响[J].中南林业科技大学学报,2007,27(6):26-33.
[13]王 森,谢碧霞,杜红岩,等.美国扁桃花器官的抗寒性[J].经济林研究,2007,25(2):19-22.
[14]王 森,杜红岩,杨绍斌,等.7个美国扁桃品种在河南南阳的引种表现[J].经济林研究,2008,26(2):65-68.
[15]Yelenosky G. Bark-splittlng from freeze injury of young citrus on different rootstocks [J].Proc Fla State Hort Soc, 1979, 92:28.
[16]常红伟.延边地区不同梨品种解剖结构与抗寒性关系的研究[D].长春:东北师范大学, 2008.
[17]王美琴.花椒枝条组织结构与抗寒性的关系[J].甘肃科学学报,2008,20(4):45-47.
[18]Mattheis J P, Ketchie D O. Changes in Parameters of the Plasmalemma ATP Palse During Cold Acclimation of Apple Tree Bark Tissues [J].Physiologia Plantarum,1990,78:616-622.
[19]彭伟秀,杨建民,张 芹,等.几个仁用杏品种枝条组织结构与抗寒性关系的初步研究[J].河北农业大学学报,2002,25(1):48-50.
[20]崔国文,马春平.紫花苜蓿叶片形态结构及其与抗寒性的关系[J].草地学报,2007,15(1):70-75.
[21]郑家基,卢 炜,陈利恒,等.龙眼、橄榄叶片空隙率与耐寒性的关系[J].福建农业大学学报,1996,(2):161-164.
[22]Chen Y R. Observation of soybean root meristematic cells in response to that heat shock[J]. Protoplasma, 1998, 144: 1-9.
[23]Lee S H, Singh A P, Chung G C,et al. Chilling root temperature causes rapid ultra structural changes in cortical cells of cucumber(Cucumis sativusL) root tips[J]. Journal of Experimental Botany,2002, 53 (378): 2225-2237.
[24]Mai A J. Fine structure of the ray parenchyma cells inPopulus tremuloidesin relation to senscence and seasonal changes [J].Tex J Sci,1979,24:245-260.
[25]Herminda Reinoso, Virginia Luna, Richard P Pharis,et al.Dormancy in peach (Prunus persica) flower buds. V. Anatomy of bud development in relation tophenological stage [J]. NRC Canada Can J Bot, 2002, 80: 656-663.
[26]房 用,李秀芬,慕宗昭,等.茶树抗寒性研究进展[J].经济林研究,2004,22(2):69-72.
[27]唐立红,赵雪梅,朱 月,等.自然降温过程中紫斑牡丹叶片结构与抗寒性关系的研究[J].北方园艺,2012(11):64-66.
[28]邹雪娟,李润唐,吴 钿,等. 3个菠萝品种叶片组织结构及其与抗寒性的关系研究[J].广东农业科学,2012,5:38-40.
