澳洲坚果花芽分化期碳水化合物含量的变化动态
2013-04-04杜丽清邹明宏陆超忠罗炼芳张汉周
曾 辉 ,杜丽清 ,邹明宏 ,陆超忠 ,罗炼芳 ,张汉周
(1.中国热带农业科学院 南亚热带作物研究所,广东 湛江 524091;2.农业部热带果树生物学重点实验室,广东 湛江 524091)
澳洲坚果Macadamia integrifolia的花期较长,花量大,但坐果率较低。花芽分化质量是影响坐果率的因素之一,营养是花芽分化及花器官形成与生长的物质基础,其中碳水化合物对花芽的形成尤为重要。已有很多报道指出,碳水化合物的积累与花芽分化密切相关[1-3]。目前,国内外有关澳洲坚果花芽分化方面的研究主要涉及其开花授粉特性、花粉活力、柱头可授性、温度对花期的影响、花芽分化期间内源激素的变化及赤霉素对成花的影响等方面[4-10],而少见有关于与澳洲坚果花芽分化密切相关的碳素营养变化动态的研究报道。为此,文中以澳洲坚果品种Own choice为试料,通过测定花芽分化期间碳水化合物的含量,探讨了澳洲坚果花芽分化期间叶片和枝条中碳水化合物含量的变化规律,以期为生产中通过人工调控来提高澳洲坚果花芽分化的质量提供借鉴。
1 材料与方法
1.1 试验地概况
试验地位于广东省湛江市南亚热带作物研究所澳洲坚果园,地处21°9′ N,110°15′ E,海拔20 m。试验地为平地,土壤为砖红壤。
1.2 试验材料及处理方式
随机选取生长良好、树体大小基本一致的5年生Own Choice澳洲坚果树为试验材料,试验植株为嫁接树,砧木采用澳洲坚果的品种Hinde(H2)的实生苗。试验共设置3个处理,每个处理各选5株树:处理①即赤霉素处理,2005年10月1日和10月10日2次对植株进行赤霉素(浓度为40 mg·L-1)喷雾处理;处理②即环割处理,2005年10月1日对植株的主枝(每株选2个直径为2.5~3.0 cm的主枝)进行螺旋环割处理,环割一圈,宽度为4~5 mm,深度到木质部;处理③即不做任何处理,用作对照。试验期间,对试验植株每周浇水1 次,如遇降雨则暂缓浇灌。
1.3 物候期的观测
在2005年9月至2006年3月和2006年9月至2007年3月这两个时间段内,每隔7 d观察记记录植株的物候变化情况,包括秋梢萌动期、展叶期、转绿期、秋梢老熟期、花穗开始伸长(花穗长达5 mm)和开花时间等。每节花穗数量的统计方法是:在每株树的北面和南面随机各选一个粗度约为3 cm的大枝,随机调查其上短枝(长10~15 cm)的节数及其着生的花穗数量。每个处理调查3株树。观察各处理花芽出现的时间、花穗开始伸长时间及盛花期,并对成花状况进行统计。
1.4 取样方法
2005年9月1日至2006年1月21日,每隔20 d取样1次。取样器官为短枝及第二轮生长的叶片,样品用塑料袋密封后放入冰壶中速冻,带回实验室洗净,将枝条、叶片样品分成2份,一份存放于-80 ℃的冰箱中以待用,用于果糖、葡萄糖和蔗糖的测定;另一份样品用牛皮纸信封装好,立即放到105 ℃的烘箱中杀青15 min,然后在65 ℃下烘干,粉碎后过80目筛,然后将样品密封置于-20 ℃的冰箱中保存,用于测定可溶性总糖和淀粉的含量。
1.5 可溶性糖的测定
可溶性糖的提取:采用烘干样品,用80%的乙醇在80 ℃下水浴提取,用苯酚显色法测定[11]。于721分光光度计485 nm下比色测定吸光值(OD值)。
1.6 淀粉的提取及测定
样品的提取与测定:提取可溶性糖的残渣,移入50 mL的容量瓶中,加入20 mL热蒸馏水中,放入沸水浴中煮沸15 min,再加入52%高氯酸3 mL提取15 min,冷却后,摇匀,并用蒸馏水定容,加入0.5 g高岭土,摇匀。加入5 mL溶液到离心管中,2 500 r·min-1离心6 min,取其上清液1 mL,再加1 mL蒸馏水,加9%苯酚溶液1 mL,摇匀至无絮状物,从液面正上方在15~20 s内加入5 mL 浓硫酸,摇匀。在室温下放置30 min,在485 nm下比色测定吸光值(OD值),计算淀粉含量。
1.7 果糖、葡萄糖、蔗糖含量的测定
文中实验参照(有所改进) Ben-Bassat 等人[12]、高娃等人[13]、唐根源等人[14]的提取及测定方法进行,具体步骤如下:称取1 g样品,加入4 mL纯水冰浴研磨,在80 ℃下水浴提取10 min后,将提取液于10 000 r·min-1转速下离心10 min,再加入4 mL纯水于残渣中,搅拌后再于10 000 r·min-1转速下离心10 min,合并上清液,定容至10 mL,取1 mL过0.22 µm的微孔滤膜后上机测定。
色谱条件:采用美国Perkin Elmer公司的高效液相色谱仪系统设备(Series 200)进行测定。其工作条件为:选用Phenomenex NH2柱(5 µm, 4.6 mm ×250 mm);流动相为乙腈∶水为70∶30(V/V);流速为1.0 mL·min-1;柱温为35 ℃;进样量20 µL;检测器采用示差检测器(RID);糖标样为美国SIGMA公司产品;所用试剂均为色谱纯。
2 结果与分析
2.1 不同处理对枝梢生长的影响
对照处理:9月至10月中旬植株枝梢处于稳定期,10月下旬秋梢开始萌动,11月上旬至11月下旬为展叶期,11月下旬至12月中旬为转绿期,12月下旬到1月下旬秋梢老熟稳定。
赤霉素处理:9月枝梢处于稳定期,10月中旬枝梢开始萌动,陆续抽生秋梢,10月下旬开始展叶,11月下旬进入转绿期,12月下旬至1月下旬枝梢老熟稳定。
环割处理:其物候期与对照处理的基本相同,只是新梢的抽生量稍少。
试验结果表明:外源赤霉素处理能促进植株的营养生长,该处理后的植株新梢可提早萌发,而环割处理对植株的营养生长略有抑制作用,表现为新梢量减少。
2.2 赤霉素处理对花期及成花量的影响
从2005年9月至2006年3月和从2006年9月至2007年3月这两个时间段内,对3个处理植株的成花情况进行了观察,结果如表1所示。由表1可知,用赤霉素处理的植株其花芽出现的时间、花穗开始伸长的时间及盛花期均明显迟于对照植株,2005~2006年分别推迟了20、16、5 d;2006~2007年则分别推迟了22、17、7 d[9]。而环割处理植株的花期与对照处理的基本一致,但环割处理的植株其花穗伸长状态及盛花情况比对照的植株更整齐。
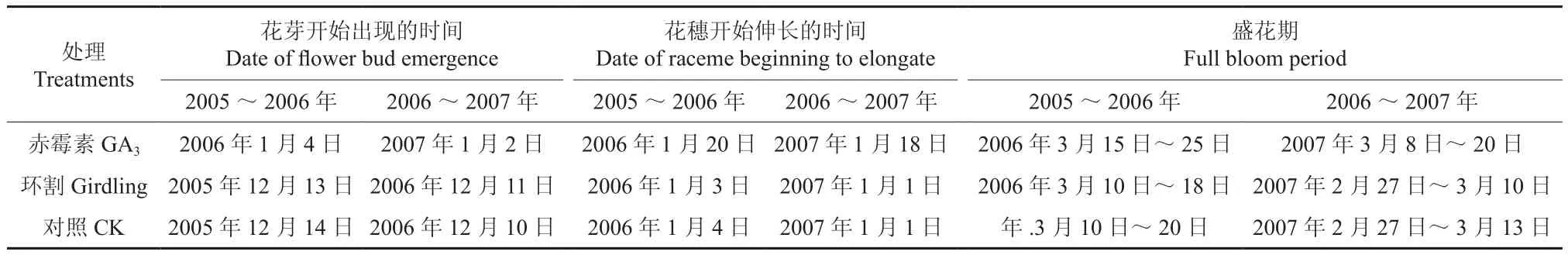
表1 2005~2007年各处理植株的花期Table 1 Flower stages of plants under different treatments from 2005 to 2007
对供试植株的成花情况进行了统计,结果如表2所示。在2005~2006年间和2006~2007年间,赤霉素处理的植株上每节花穗的数量分别为1.45和1.52穗[9],环割处理的植株上每节花穗的数量分别为2.65和2.61穗;而对照植株的每节花穗数量则分别为2.60和2.55穗。表2表明,外源赤霉素处理明显推迟了植株的花期,显著降低了每节位花穗数,此结果证明了赤霉素不利于澳洲坚果花芽的分化;环割处理植株的花期与对照处理的基本相同,每节位的花穗数量略有增加,但差异不显著。
2.3 叶片和枝条中碳水化合物含量的变化
2.3.1 可溶性糖含量的变化
花芽分化期间澳洲坚果树叶片、枝条中可溶性糖含量的变化情况如图1所示。
由图1可知,不同处理后植株叶片中可溶性糖含量的变化动态各不相同:对照植株从9月上旬开始呈上升趋势,10月中旬出现一个小的峰值(达8.23%)后下降,11月上旬以后再次上升,于12月中旬达到最大值(9.2%),此后再次下降;赤霉素处理的植株从9月上旬逐渐上升,12月中旬达到最大值(8.74%),此后缓慢下降;环割处理植株其含量变化的趋势与赤霉素处理的相似,其峰值(达9.32%)出现的时间提前到11月下旬,此后呈下降趋势,1月下旬略有回升。
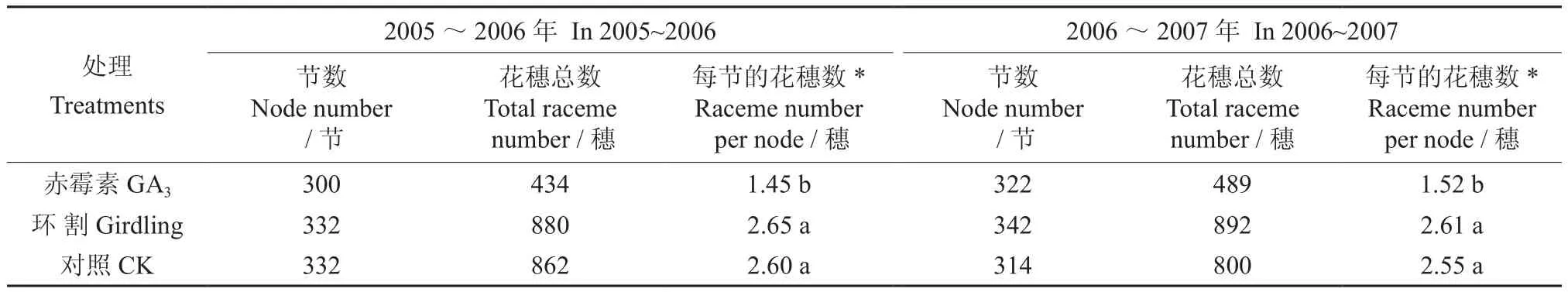
表2 2005~2007年各处理植株的成花量†Table 2 Raceme number in plants under different treatments from 2005 to 2007
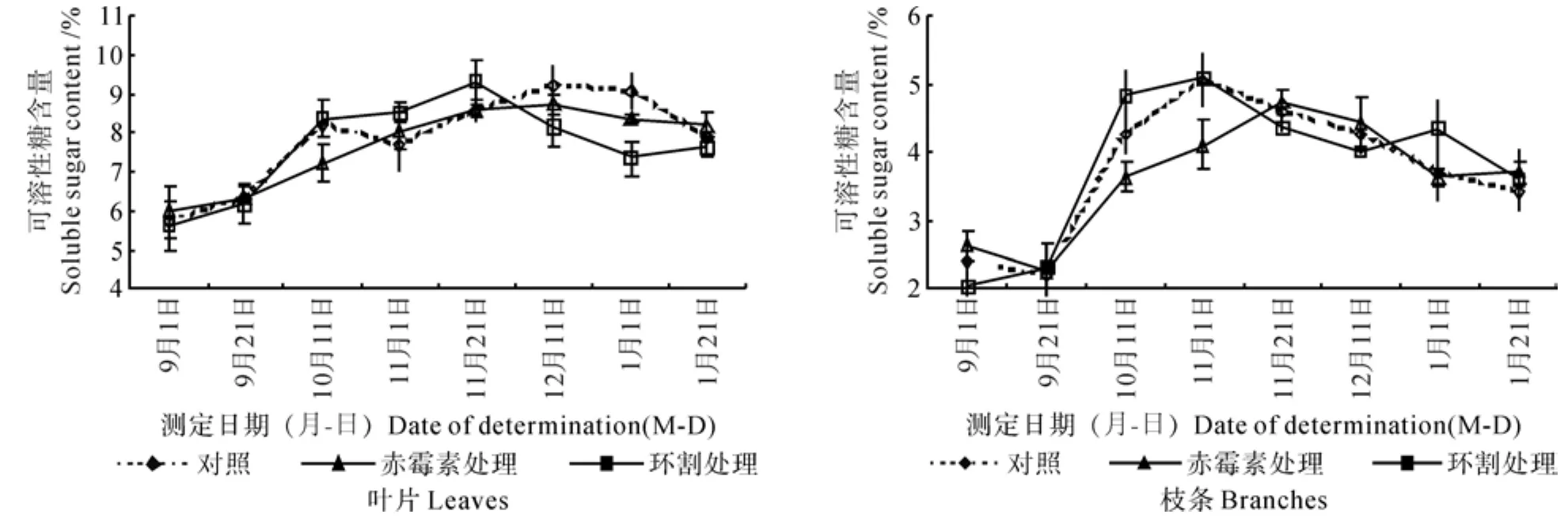
图1 花芽分化期间各种处理的叶片和枝条中可溶性糖含量的变化Fig.1 Changes of soluble sugar contents in leaves and shoots under different treatments during flower bud differentiation period
从图1中可以看出,3种处理方式下枝条中可溶性糖含量的变化趋势基本相同,9月上旬都保持在2.00%~2.50%之间;对照植株11月上旬达到最大值(5.03%),然后呈下降趋势;赤霉素处理植株的峰值出现的时间推迟到11月下旬,且峰值(4.72%)较低,9月下旬至11月下旬,其枝条中可溶性糖的含量明显低于对照;环割处理植株与对照的变化曲线相似,但9月下旬至11月上旬,其枝条中可溶性糖的含量明显高于对照。
2.3.2 淀粉含量的变化
花芽分化期间澳洲坚果树叶片、枝条中可溶性糖含量的变化情况如图2所示。
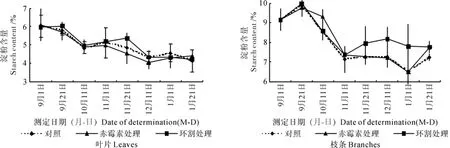
图2 花芽分化期间各处理叶片和枝条中淀粉含量的变化Fig.2 Changes of starch contents in leaves and shoots under different treatments during flower bud differentiation period
各处理植株叶片中淀粉含量的变化曲线相同,从9月上旬的6%逐步下降到4%,赤霉素处理的含量略低于对照,而环割处理的含量则略高于对照。
各处理植株枝条中淀粉含量的变化曲线相同,赤霉素处理的含量与对照基本相同,而环割处理的含量则大于对照;环割处理植株中淀粉含量在10月中旬后较高。
从器官的分布来看,叶片中可溶性糖含量高于枝条,为5.5%~9.2%,而枝条为2%~5%;淀粉含量则以枝条为高,枝条为6.5%~12.5%,而叶片中的含量则为4%~6%。
由图1和图2可知,在10月上、中旬,枝条、叶片中可溶性糖的含量急剧上升,且以环割处理后的植株糖含量上升的幅度最大,其次是对照植株,赤霉素处理植株上升的幅度稍小;叶片与枝条中淀粉的含量则呈下降趋势。
结合物候期观察来分析,从10月上旬开始,各处理的植株新梢陆续开始萌动,叶片与枝条中可溶性糖含量增加,淀粉含量则下降。
11月下旬到12月中旬,叶片进入转色期的澳洲坚果可溶性糖含量均下降,随着叶片和枝梢的老化,各处理植株器官中淀粉的含量略有上升。
2.3.3 果糖、葡萄糖和蔗糖含量的变化
2.3.3.1 果糖含量的变化
花芽分化期间澳洲坚果树叶片、枝条中果糖含量的变化情况如图3所示。
由图3可知,对照处理植株叶片中果糖含量的变化情况是先下降,然后迅速上升,11月下旬达到最大值,然后迅速下降,12月中旬后又有所回升。环割处理植株叶片中果糖含量的变化情况与对照的基本相同,而赤霉素处理植株叶片中的果糖含量,9月下旬就开始回升,9月上旬至下旬,其含量与对照的相同,9月下旬至11月上旬,其含量高于对照,之后其含量又低于对照。
各处理植株枝条中果糖含量的变化趋势与叶片略有不同。对照处理植株枝条中果糖含量,9月上中旬急剧上升,从9月上旬的400~500 µg·g-1上升到700~750 µg·g-1,11月上旬各处理均降到最低值(90~300 µg·g-1),11月下旬迅速上升,达到500~700 µg·g-1,此后迅速下降到9月上旬的水平。与对照比较,赤霉素处理植株枝条中果糖含量较高,环割处理植株枝条中果糖含量除了11月上旬略高外,其它时期与对照无明显差异。
2.3.3.2 葡萄糖含量的变化
花芽分化期间澳洲坚果树叶片、枝条中葡萄糖含量的变化情况如图4所示。
由图4可知,9月上旬至9月下旬,各处理植株枝条及叶片中葡萄糖含量变化不大,枝条一般保持在700 µg·g-1左右,而叶片在900~1 300µg·g-1之间,11月下旬均达到最大值,枝条在1 300~1 600 µg·g-1之间,叶片在1 800~2 300µg·g-1之间,此后各处理枝条与叶片中葡萄糖含量略有下降。与对照相比,赤霉素处理植株叶片中葡萄糖含量无明显差异,枝条中葡萄糖含量在10月中旬明显较低,而在11月下旬则较高,其它时期无明显差异;环割处理植株叶片中葡萄糖含量在10月上旬到12月上旬略高,枝条中葡萄糖含量在11月中旬以后较低,其它时期无明显差异。
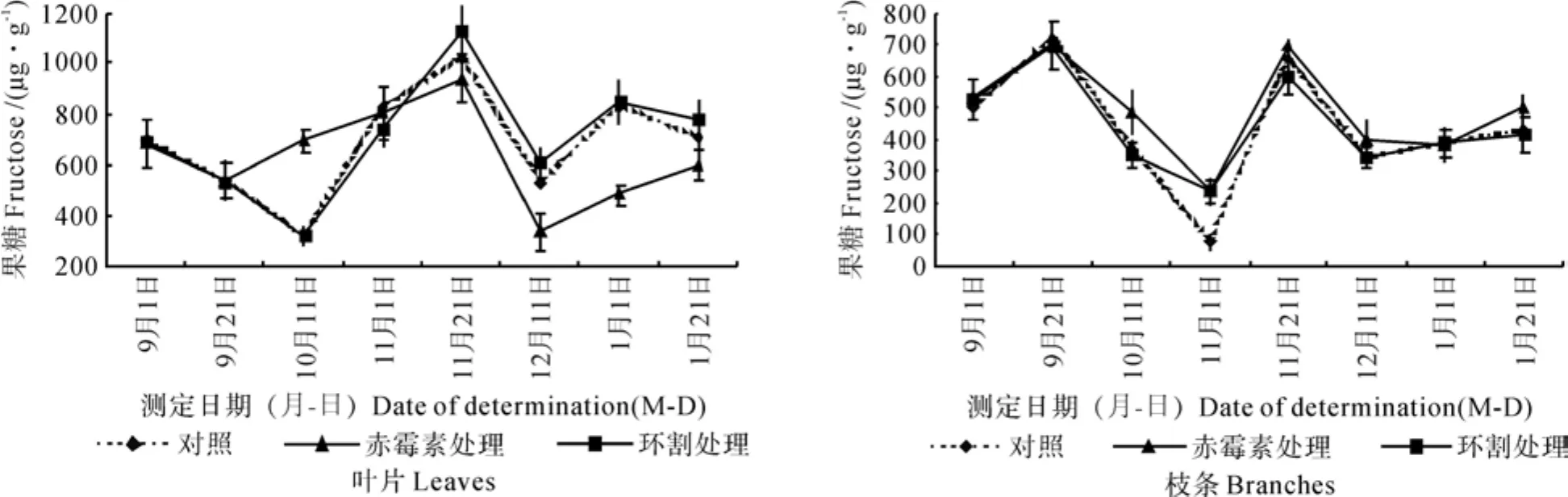
图3 花芽分化期间各处理叶片和枝条中果糖含量的变化情况Fig.3 Changes of fructose contents in leaves and shoots under different treatments during flower bud differentiation period

图4 花芽分化期间各处理叶片和枝条中葡萄糖含量的变化情况Fig.4 Changes of glucose contents in leaves and shoots under different treatments during flower bud differentiation period
2.3.3.3 蔗糖含量的变化
花芽分化期间澳洲坚果树叶片、枝条中蔗糖含量的变化情况如图5所示。

图5 花芽分化期间各处理叶片和枝条中蔗糖含量的变化情况Fig.5 Changes of sucrose contents in leaves and shoots under different treatments during flower bud differentiation period
9月上旬到下旬,各处理各器官蔗糖含量变化不大,叶片一般保持在600 µg·g-1左右,枝条在200 µg·g-1左右,11月下旬枝条和叶片中蔗糖含量达到最大值,叶片在800~1 100 µg·g-1之间,枝条在500~650 µg·g-1之间,此后又有一定幅度的下降。与对照比较,赤霉素处理植株枝条中蔗糖较低(11月下旬略高),而叶片中蔗糖较高(9月上中旬略低),环割处理植株与对照无明显差异。
从图4和图5可知,从10月上旬至11月下旬,经各种处理后澳洲坚果树的枝条和叶片中葡萄糖和蔗糖的含量呈明显上升趋势,此后则稍有回落。
综上所述,花芽分化期间澳洲坚果的枝条和叶片中有较高浓度的可溶性糖含量和较低浓度的淀粉含量,可溶性糖中又以葡萄糖和蔗糖含量较高。
3 讨 论
3.1 梢期对花芽分化的影响
澳洲坚果的花期较长,花期长短与当地的气候条件有关。在夏威夷,花期长达14周以上[7],在广东湛江则花期比较集中,一般为2~4周,而在云南德宏地区的花期稍长,全年都有花出现,但主要集中在3月至4月。澳洲坚果的成花枝一般是树冠内膛1.5~3年生的短枝,花芽在叶腋处或叶片脱落处形成。从表1可以看出,2006年经赤霉素处理的植株花穗开始伸长及盛花期分别比对照晚16 d和5 d。从物候观察的结果可知,经赤霉素喷布处理的植株在10月中旬秋梢开始萌动,而经环割处理的植株和对照植株在10月下旬才开始萌动。结果表明,秋梢萌动早,花穗开始伸长及盛花期晚。这可能是由于秋梢萌动导致树体内GA3、IAA等含量升高,进而使花芽分化受到抑制的缘故。
3.2 碳水化合物对花芽分化的影响
营养生长和营养物质的积累是花芽分化的物质基础,花芽的分化和发育是一个形态建成过程,需要大量的营养物质来供应完成,碳水化合物对花芽的形成有非常重要的作用[1]。淀粉的积累对成花有利[3],对花芽形成的质量起重要作用[2]。梁芳等人[15]试验发现,在花芽分化过程中,芽中的可溶性总糖、蔗糖和淀粉含量迅速增加,表明高含量的碳水化合物有利于菊花花芽分化。陈厚彬等人[16]发现,花发端前,荔枝高成花率树的淀粉含量以小枝最高,形成从小枝到末端秋梢和叶片的下降梯度,而低成花率树则没有这种梯度。李静等人[17]芍药花芽分化主要靠可溶性糖提供营养物质,在可溶性糖浓度降低到一定水平之后,淀粉水解以供花芽分化过程中对营养物质的需求。营养物质的供应是否充足,决定花芽分化质量的优劣[2]。叶片中可溶性糖的积累有利于草莓、奈李的花芽分化[18-19]。杨义标等人[20]报道,勒杜鹃花芽分化过程中可溶性糖含量先升后降。吴月燕等人[21]研究了葡萄叶片中碳水化合物的变化对花芽分化的影响,结果表明,花芽分化进度与可溶性糖、蔗糖含量呈极显著正相关,与果糖含量呈显著正相关,叶片中的淀粉积累有利于花芽分化,叶片的淀粉含量与花芽分化呈显著正相关。郭金丽等人[3]报道,苹果梨花芽生理分化期,成花短枝和叶片中淀粉积累快,形态分化期,成花短枝中淀粉大量积累。成花诱导并不一定要求很高的碳水化合物水平,而是强调其“可利用性”。
文中的试验结果表明,在澳洲坚果花芽分化期间,枝条与叶片中可溶性糖含量持续上升,并处于一个较高的水平,且花芽萌动早的对照和环割处理比花芽萌动迟的赤霉素处理要高,表明可溶性糖的积累有利于澳洲坚果花芽分化,这与前人的研究相一致。在可溶性糖中,以葡萄糖和蔗糖浓度较高。本试验中,在花芽诱导后期枝条和叶片中淀粉含量均有所下降,表明枝条及叶片中可能存在淀粉向可溶性糖转化的过程,这一过程可能有助于增加树体中碳水化合物的可利用程度,促进澳洲坚果的花芽分化。
参考文献:
[1] 马焕普.果树花芽分化与激素的关系[J].植物生理学通讯,1987, (1):1 - 6.
[2] 李天红,黄 阵.苹果花孕育机理的探讨[J].植物生理学报,1996, 22 (3):251 - 257.
[3] 郭金丽,张玉兰.苹果梨花芽分化期蛋白质、淀粉代谢的研究[J].内蒙古农牧学院学报,1999,20(2): 80-82.
[4] 陶 丽,贺熙勇,倪书邦,等.澳洲坚果开花授粉特性的研究[J].热带作物学报,2010, 31(3):349-354.
[5] 陶 丽,倪书邦,贺熙勇,等.不同贮藏方式对澳洲坚果花粉萌发率的影响[J].中国南方果树,2010,39 (3):52-54.
[6] 曾黎明,陈显国,林玉虹,等.澳洲坚果花粉活力、柱头可授性比较研究[J].江苏农业科学,2012, 40 (3): 123-125.
[7] Nakata S. Progress report on flowering, nut setting and harvesting,with special reference to the effects of night temperatures and growth regulators [J]. Proc Hawaii Macadamia Prod Assoc, 16th Annu Meet, 1976, 5:31 - 36.
[8] Moncur M W, Stephenson R A, Trochoulais T. Floral development ofMacadamia integrifoliaMaiden & Betche under Australia conditions[J]. Scientia Hortic, 1985,(27): 87 - 96.
[9] 曾 辉,陈厚彬,杜丽清,等.喷施赤霉素推迟和减少澳洲坚果成花的效应[J].果树学报2008,25(2):203-208.
[10] 曾 辉,杜丽清,邹明宏,等.澳洲坚果花芽分化期间内源激素的变化[J].安徽农业科学, 2008,36(34):14949-14953.
[11] 向曙光,刘思俭,朱万洲,等.苯酚法测定植物组织中的碳水化合物[J].植物生理学通讯,1984,19(2): 42-44.
[12] Ben-Bassat A A, Grushka E. High performance liquid Chromatography of mono-and oligosaccharide[J]. Journal of Chromatography, 1991, 14(6):1051.
[13] 高 娃.高压液相色谱分析常见单糖、多糖[J].生物技术,1998, 8(2):44 - 46.
[14] 唐根源.高效液相色谱法测定萘果中的山梨醇和糖类[J].色谱 , 2000,18(5):459 - 461.
[15] 梁 芳,郑成淑,张翠华,等.菊花花芽分化过程中芽和叶片碳水化合物含量的变化[J].山东农业科学,2008,(1): 40-42.
[16] 陈厚彬,黄辉白,刘宗莉.荔枝树成花与碳水化合物器官分布的关系研究[J].园艺学报,2004,31(1): 1-6.
[17] 李 静,郭绍霞.芍药花芽分化期间营养物质与酶活性的变化[J].北方园艺, 2011,(7):73-75.
[18] 钟晓红,罗先实,陈爱华.奈李花芽分化与体内主要代谢产物含量的关系. 湖南农业大学学报, 1999, 25(1):31-35.
[19] 孙乃波,张志宏.草莓花芽分化过程中叶片碳水化合物和蛋白质含量的变化[J].安徽农业科学, 2006,34(1):2328-2329.
[20] 杨义标,邵志芳,邱少松,等.勒杜鹃花芽分化过程中内源激素含量和碳氮营养的变化研究[J].中国农学通报, 2008,24(2):281 - 284.
[21] 吴月燕,李培民,吴秋峰.葡萄叶片内碳水化合物及蛋白质代谢对花芽分化的影响[J].浙江万里学院学报,2002,15(4):54-57.
