“金丝4号”枣大小孢子发生及雌雄配子体发育
2013-04-04袁德义段经华冯延芝
邹 锋 ,袁德义 ,段经华 ,廖 婷 ,冯延芝 ,高 超
(1.中南林业科技大学 经济林育种与栽培国家林业局重点实验室,湖南 长沙 410004;2.中国林业科学研究院,北京 100091)
“金丝4号”枣Ziziphus jujubeMill cv ‘jinsi No. 4’原产于山西运城龙居乡东辛庄一带,是从“金丝2号”自然杂交的实生枣树中选育而来,为枣树中稀有的名贵鲜食品种[1]。“金丝4号”耐瘠薄,适应性强,自19世纪80年代对枣进行良种选育和资源开发利用以来,“金丝4号”已逐步推广到全国十余个省、市及地区。“金丝4号”以早实、果实特大、皮薄肉厚、清香甜脆、风味独特著称,受到人们的重视和消费者的欢迎。其鲜果含糖量22.75%,含酸量0.368%,含Vc 2.92 mg·g-1,可食率达 97.3%,营养丰富,品质上等。
枣树在林业产业结构调整、山区经济发展和林产品外贸出口中占有特殊重要的地位,被称为“木本粮食”、“铁杆庄稼”[2]。枣在生产中普遍存在结实率低、产量低的现象,这严重影响着枣的经济效益及其产业的发展。目前,关于枣的研究多集中在引种栽培[3]、果实品质[4-5]、病虫害防治[6]、遗传多样性[7]等方面,而关于枣结实率低的生殖生物学因素的研究报道较少[8-13]。植物的有性生殖过程是植物体进行物种延续和种群扩大的根本动力。开花、传粉、受精、结实,是一个连续的过程,只有从整体的和发育的角度去研究探讨,才能找出问题所在。而大、小孢子发生及雌、雄配子体的发育状况是果树授粉受精和果实产量形成的基础。因此,本研究中试图通过对“金丝4号”枣大小孢子发生及雌雄配子体发育过程进行观察,为揭示枣结实率低的内在原因提供理论依据,为提高枣产量和品质以及开展杂交育种研究奠定基础。
1 材料与方法
试验材料采自中南林业科技大学衡阳枣种质资源圃,试验地位于东经 112°18′54″、北纬26°54′48″、海拔70.0 m的缓坡丘陵红壤。试验地水肥条件良好,采取常规管理方式。
选取正常开花结果的8年生“金丝4号”3株,从2010~2011年每年的4~6月,每3天采样1次,每次从采样树冠中部的不同枣吊的基部选取大小发育情况较为一致的“金丝4号”花芽或花蕾10个,用FAA(70%酒精90 mL、醋酸5 mL、福尔马林5 mL)固定、抽气,后转入70%酒精置于4 ℃冰箱保存备用。石蜡切片方法参考袁德义等[14]的方法,铁钒-苏木精染色,曙红对染,加拿大中性树胶封片,OLYMPUS BX-51显微镜下观察照相。
2 结果与分析
2.1 花药壁的发育
从“金丝4号”枣花芽的石蜡切片中可以观察其花药壁的发育过程。首先,形成枣花药原始体;花药原始体随后形成孢原细胞(见图1A),孢原细胞的典型特征是原生质浓厚,形成的场所位于花药四角;孢原细胞形成之后,通过平周分裂的方式形成2层细胞;初生造孢细胞随即形成(见图1B),即孢原细胞分裂而成的2层细胞中里面的1层细胞;外面的1层为初生壁细胞,它经过数次分裂后形成花药壁,花药壁形成时为小孢子母细胞时期,花药壁从结构上分为4层,从内至外分别是中层、绒毡层、药室内壁、表皮,次生造孢细胞位于绒毡层内 (见图1C)。另外,绒毡层细胞和中层会随着发育进程而发生变化。中层在小孢子母细胞开始减数分裂时染色变深开始退化,到“金丝4号”枣花粉成熟时只留下残迹。在“金丝4号”枣花药壁发育过程中,绒毡层内含有丰富的营养物质,能够为小孢子的发育奠定充足的物质基础,因此在花药整个发育过程中表现得最为活跃,这与何春燕[15]的研究结果类似。绒毡层在“金丝4号”枣小孢子母细胞处于减数分裂时,其开始径向加厚(见图1D);在“金丝4号”枣进入小孢子四分体时期,它开始出现原位解体现象(见图1F);绒毡层在“金丝4号”枣单核小孢子时期,会继续解体,结果导致与花药壁的中层剥离(见图1J、K);当“金丝4号”枣花粉趋于成熟时,绒毡层开始溶解、退化,直至消失,“金丝4号”枣花药壁由原来的4层只剩下2层,即药室内壁和表皮(见图1O),根据Davis[16]对被子植物花药壁发育方式的分类,花药壁发育类型分为简化型、基本型、双子叶型和单子叶型4种,“金丝4号”枣花药壁的发育属于基本型。Davis[16]根据绒毡层发育后期的形态差别,将绒毡层分为腺质绒毡层和变形绒毡层这2种类型[17],由于“金丝4号”枣绒毡层的退化、溶解发生之后,因其发生场所位于原位上,所以其核、质也在原位溶解,并没有发生位置上的移动,从而决定着“金丝4号”枣的绒毡层属于前一种类型,即为腺质型绒毡层,这与攸县油茶[18]的绒毡层类似。
2.2 小孢子发生及雄配子体发育
2.2.1 小孢子发生
从花药的横切面上看,“金丝4号”的花药具有4个花粉囊,中间有药隔隔开,整体形状呈蝴蝶状(见图1C)。由此可见,对于“金丝4号”的幼嫩花药而言,其结构组成包括初生壁细胞和造孢细胞 (见图1A、B)。对于造孢细胞,在其发育过程中,会有多次的有丝分裂,并分化形成小孢子母细胞,造孢细胞的特点是细胞体积较大、细胞核较大、细胞质浓厚、无明显的液泡;初生壁细胞在经过平周分裂和垂周分裂后形成3个细胞层,即药室内壁、中层、绒毡层,并与表皮一起组成花药壁(见图1C)。小孢子母细胞进行2次减数分裂,如减数Ⅰ末期(见图1D),减数Ⅱ末期(见图1E),其胞质分裂为同时型,经过2个连续的减数分裂过程后进入四分体时期(见图1F)。在四分体时期,4个小孢子共处于1个胼胝质(见图1F)的小孢子母细胞壁内,小孢子四分体为四面体形。
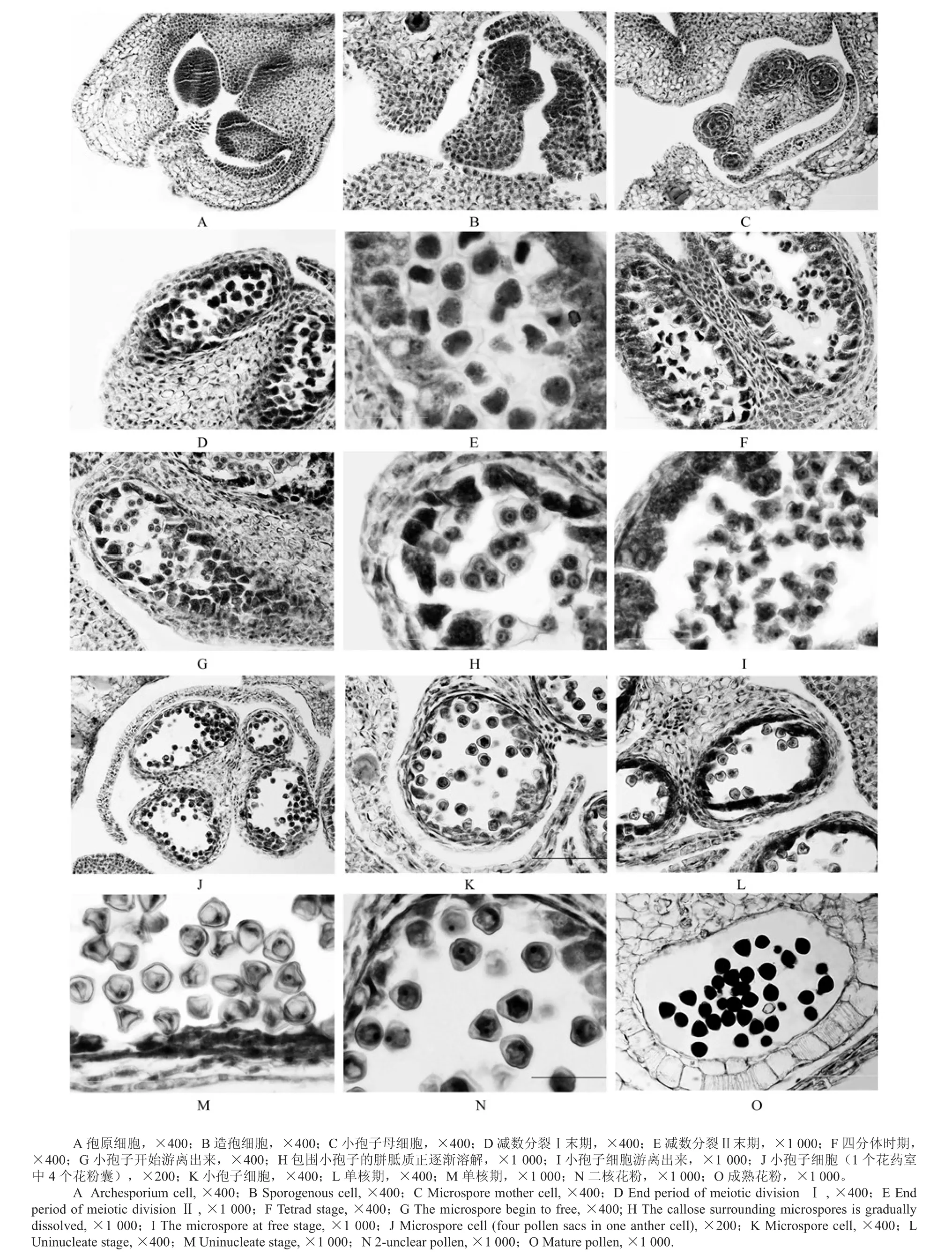
图1 “金丝4号”枣小孢子和雄配子体的形成和发育Fig.1 Formation and development of microspore and male gametophyte in Ziziphus jujube Mill cv ‘jinsi No. 4’
2.2.2 雄配子体发育
“金丝4号”枣雄配子体发育是从小孢子开始的,随着小孢子四分体之间的胼胝开始解体、溶解并消失(见图1G、H),其小孢子逐渐从四分体中游离出来(见图1I),形成单核小孢子(见图1J),这标志着其小孢子进入了雄配子体的发育阶段,雄配子体阶段对于小孢子的发育意义重大,小孢子在从四分体中刚释放出来时,细胞壁较薄,细胞质浓厚,细胞核处于细胞中央,细胞的体积总体上还比较小(见图1J)。小孢子在发育过程中从其周围的组织或器官中汲取营养成分,从而使得花粉粒体积不断增大(见图1K),并发育形成核物质(见图1L),同时小孢子体积继续增大,细胞质也明显液泡化,此时形成单核时期小孢子(见图1M)。小孢子经过一段时期生长发育就会形成3个萌发孔,萌发孔位于小孢子的表面,是供给小孢子营养物质的通道,营养物质的来源是绒毡层(见图1M)。随着小孢子不断地吸收绒毡层的水分和养分,其细胞体积增大,细胞壁加厚并且细胞质液泡化加剧,接着花粉粒的体积明显增大(见图1M),逐渐发育成为二核花粉(见图1N),并充满淀粉、脂肪等营养物质,最终花粉粒发育成熟(见图1O)。“金丝4号”花粉为三角形,具3个萌发孔,3沟,成熟花粉为2-细胞型。
2.3 大孢子发生与雌配子体发育
2.3.1 大孢子发生
根据“金丝4号”枣幼嫩花芽横切面观察,胚珠原基发生于子房壁的胎座上(见图2A),孢原细胞由珠心表皮下2~3层细胞分化而来,接着内外珠被伸长发育形成环状突起(见图2B)。紧贴珠心表皮分化出来的孢原细胞与相邻细胞相比体积明显偏大,核大而明显,细胞质浓,直接行使着大孢子母细胞的功能(见图2C)。大孢子母细胞开始进行减数分裂,经过中期(见图2D)、后期(见图2E)、末期(见图2F),胞质分裂为横向,形成四分大孢子(见图2G),靠近珠孔端的3个细胞退化,合点端功能大孢子体积增大,继续进行减数分裂发育形成单核胚囊(见图2H)。
2.3.2 雌配子体发育
单核胚囊为雌配子体发育的起点,此时其内外珠被已形成。大孢子母细胞进行减数分裂后形成四分体大孢子,四分体大孢子的排列是以直线型的方式(见图2G),其中功能大孢子位于合点端,二核胚囊的形成源于单核胚囊的1次有丝分裂,2个核由于液泡的推移作用分别移向两极(见图2I),并再各进行1次有丝分裂形成四核胚囊(见图2J),形成四核胚囊后逐渐拉长直至出现大液泡化,随后形成八核胚囊(见图2K),途径仍是通过每个核继续进行的1次有丝分裂。八核胚囊长期存在,因此断定“金丝4号”枣的成熟胚囊的完整形态:1个卵细胞、2个助细胞(其珠孔端有大液泡)、2个极核、合点端有3个反足细胞(见图2L~O)。胚囊形成过程表明“金丝4号”枣为倒生胚珠,双珠被,厚珠心,胚囊结构为蓼型。
3 结论与讨论
3.1 “金丝4号”枣胚胎学特征
“金丝4号”枣的胚胎学特征与已报道的长红枣[9]、宁夏圆枣[10]、灰枣[13]、酸枣[19]等基本一致。“金丝4号”枣的花药4室,花药壁4~5层,分别为表皮、1~2层药室内壁、中层和腺质绒毡层,花药壁的发育类型属于基本型;小孢子母细胞经2次减数分裂后形成四分体,其孢质分裂为同时型,四分体排列多为正四面体;成熟花粉为2-细胞型,其花粉为三角形,具3个萌发孔,3沟。“金丝4号”枣雄配子体发育过程中未发现大量花粉败育的现象,说明“金丝4号”枣花粉雄性可育,这不是枣子结实率低的原因。倒生胚珠,双珠被,其珠心类型为厚珠心;大孢子母细胞减数分裂形成4个大孢子,呈线形排列,合点端大孢子发育成为功能大孢子,功能大孢子经有丝分裂后形成成熟胚囊具7细胞8核,胚囊发育为蓼型。但是,有关枣胚囊发育的类型,仍存在争议。本研究结果显示,“金丝4号”枣胚囊的发育类型为蓼型,这与Srinivasachar[20]的观察结果一致,但是Kaiale[21]、田惠桥[10]认为枣胚囊发育类型是葱型,这种差异的产生可能与不同枣品种的遗传特性有关,具体原因还有待于进一步研究。
3.2 “金丝4号”枣雌雄异熟现象
植物雌雄异熟性是指1株植株或1朵花中的雌蕊和雄蕊成熟时间不一致或雌雄性功能在时间上的分离[22]。有关学者经研究推测,小孢子母细胞在结构上因有原生质道相连,形成了合孢体,从而在营养运转、分配等方面形成一个完整的系统,因此认为小孢子母细胞减数分裂是同步完成的[17],而本研究结果显示“金丝4号” 枣小孢子母细胞减数分裂不同步,这与油茶[14]类似,分析其原因,一方面可能是品种自身的特性所导致,目前尚未知其它枣品种是否也存在此现象;另一方面可能是营养因素所导致,即胼胝质壁形成过早而堵塞细胞质道,养分的运转和分配也由此受到了影响。“金丝4号”枣在5月下旬至6月初开花,开花数量多,花期较长,气温较高,传粉
昆虫较多,这非常有利于其传粉受精和坐果。高等显花植物小孢子母细胞减数分裂大量不同步现象及不同药室、不同雄蕊的发育不同步现象等均是一种有效的延长传粉期的生殖机制,即雌雄异熟机制[18]。枣为雌雄同株植物,“金丝4号”枣在其雌雄蕊发育时期存在不同步现象,即雌雄异熟,“金丝4号”枣的这种雌雄异熟现象主要表现在雄蕊的发育先于胚囊的形成,这种类型的雌雄异熟现象在油茶[15]和梨[23]上也有发现。由此说明“金丝4号”枣是异花授粉或以异花授粉为主的植物,其雌雄配子体的形成和发育过程正常,这种雌雄异熟机制在植物界中普遍存在,其目的是促进异交或远交,限制自交或近交所导致的物种衰退或退化[23-24]。
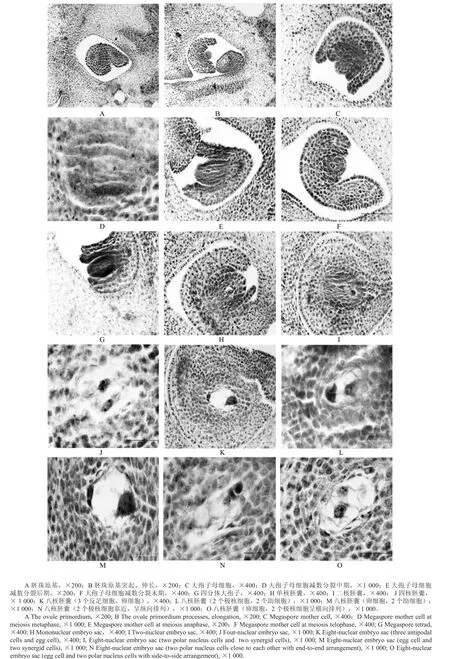
图2 “金丝4号”枣大孢子和雌配子体的形成和发育Fig.2 Formation and development of megaspores and female gametophyte in Ziziphus jujube Mill cv ‘jinsi No. 4’
参考文献:
[1] 王奎武,韩振虎,刘海波,等. 金丝4号枣引种栽培试验初报[J]. 林业科技 ,2002,27(4):54 - 56.
[2] 曲泽州,王永蕙. 中国果树志·枣卷[M]. 北京:中国林业出版社,1991.
[3] 樊保国,李月梅,李登科.冬枣引种栽培区气候适宜性的灰色综合评估[J]. 中国农学通报,2011,27(8):208-211.
[4] 张 萍,史彦江,宋锋惠,等.南疆灰枣主要营养品质性状的变异及相关性研究[J]. 果树学报,2011,28(1):77-81.
[5] 卢桂宾,李春燕,郭晓东. 外源钙肥对枣果实矿质营养元素含量的影响[J]. 经济林研究,2010,28(3):69-74.
[6] 侯晓杰. 枣缩果病病原和防治研究[D]. 保定:河北农业大学,2010.
[7] 徐呈祥. 等渗透势干旱、盐、碱胁迫下5个枣品种及酸枣的生物学响应与抗逆性[J]. 中南林业科技大学学报,2012,32(12): 139 - 146.
[8] 王勋陵. 枣果实发育解剖学的初步观察[J]. 植物学报,1974,16(2): 161 - 179.
[9] 王群瑛. 枣胚与胚乳的发育[J]. 植物学报,1983,25(6):526-531.
[10] 田惠桥,马德兹. 一种单性结实枣的胚胎学观察[J]. 植物学报 ,1987,29(1):29 - 33.
[11] 祁业凤. 枣胚败育机理与胚培养研究[D]. 保定:河北农业大学,2002.
[12] 王玖瑞. 枣树雄性不育和胚败育研究[D]. 保定:河北农业大学,2004.
[13] 张学英,彭士琪,郭振怀. 枣授粉受精及胚胎发育研究[J]. 林业科学,2004,40(5):210- 213.
[14] 袁德义,邹 锋,谭晓风,等. 油茶花芽分化及雌雄配子体研究[J]. 中南林业科技大学学报,2011,31(3):65-70.
[15] 何春燕. 普通油茶有性生殖过程研究[D]. 长沙:中南林业科技大学,2009.
[16] Davis G L. Systematic Embryology of the Angiosperms [M].New York: John Wiley & Sons, 1966.
[17] 胡适宜. 被子植物生殖生物学[M]. 北京:高等教育出版社,2005.
[18] 邹 锋. 攸县油茶生殖生物学研究[D]. 长沙:中南林业科技大学,2010.
[19] 牛瑜菲,彭建营,李 莉. 枣及酸枣小孢子不同发育时期的花器形态与解剖特性研究[J]. 植物遗传资源学报,2011,12(1):158-162.
[20] Srinivasachar D. Embryological studies of some member of Rhanaceae[J]. Proc India Acad B, 1940, 11:107 - 116.
[21] Kajale LB. A contribution to the life-history ofZiziphus jujubeLamk[J]. Proc Nat Sci India, 1944,10(4): 387 - 390.
[22] 赵书岗,王红霞,张志华. 核桃雌雄异熟性研究进展[J]. 湖北农业科学,2011,50(17):3466-3470,3475.
[23] 段经华. 早生黄金梨授粉受精生物学研究[D]. 长沙:中南林业科技大学,2006.
[24] 张大勇. 植物生活史进化与繁殖生态学[M]. 北京:科学出版社,2003.
