过表达小鼠UNCV蛋白质对HeLa细胞凋亡的影响
2013-02-05刘冰徐嫄赵爽王冬平杨蕾蕾李微尚世臣张广州李文龙曾林
刘冰,徐嫄,赵爽,王冬平,杨蕾蕾,李微,尚世臣,张广州,李文龙* ,曾林*
(1.军事医学科学院实验动物中心,北京 10071;2.军事医学科学院放射与辐射医学研究所,北京 100850)
Uncv 无毛小鼠是军事医学科学院实验动物中心培育的来源于BALB/c 品系的自发性被毛缺失突变小鼠。纯合突变小鼠皮肤无毛,杂合突变小鼠皮肤背毛稀疏[1-2]。研究表明,该小鼠无毛突变性状是由常染色体单基因突变所引起,通过基因连锁分析,最终确定导致无毛性状的突变基因,并命名为Uncv[3]。因此对该基因生物学功能的研究成为亟待解决的问题。本实验室已经克隆出小鼠Uncv 基因并成功构建真核表达载体[4]。
细胞凋亡是指细胞的程序性死亡,即细胞在一定的生理或病理状态下,由自身基因调控的一种特定的主动性死亡过程。细胞凋亡在生物体的发育成熟、进化以及细胞功能维持等方面具有重要的生物学意义。肿瘤的发生不仅与细胞的异常增殖和分化有关,也与细胞凋亡的异常有关,恶性肿瘤的自发凋亡也会起到治疗肿瘤的作用[5]。临床上治疗肿瘤的主要手段是利用化疗或者放疗促使肿瘤细胞凋亡增加。在对Uncv 基因的研究过程中,我们发现它能影响与肿瘤发生和发展相关的信号通路,那么UNCV 蛋白质是否能影响宫颈癌细胞株(HeLa)的凋亡呢?
本研究采用脂质体转染和G418 筛选的方法获得稳定过表达UNCV 蛋白质的HeLa 细胞株。通过血清饥饿和阿霉素(doxorubicin,DOX)诱导HeLa 细胞凋亡实验,证实了过表达UNCV 蛋白质对血清饥饿和阿霉素诱导的HeLa 细胞凋亡的影响。
1 材料与方法
1.1 材料与主要试剂
本实验室已构建小鼠Uncv 真核表达质粒(pcDNA3.1-Flag-Uncv)。HeLa 细胞和真核表达载体pcDNA3.1-Flag-vector 由本实验室保存;胰蛋白酶、进口胎牛血清、脂质体转染试剂Lipofectamine 2000(均为Invitrogen 公司,美国);Western blot 发光检测试剂盒(英格恩公司,中国);凋亡检测试剂盒Annexin V-FITC(凯基公司,中国);蛋白质预染marker(Fermentas 公司,美国);Anti-Flag 单抗(Sigma 公司,德国);内参抗体beta-actin antibody(Abcam 公司,美国);HRP 标记羊抗小鼠IgG(中杉金桥有限公司,中国);DMEM/high glucose 培养基(HyClone 公司,美国)。
1.2 方法
1.2.1 稳定过表达UNCV 蛋白质HeLa 细胞株的获得
在转染前24 h,对HeLa 细胞进行定量培养,以每孔5 ×105的细胞数接种6 孔板,每孔培养液的量为2 mL。转染前1 h 将细胞培养基换至1 mL,将Uncv 重组质粒pcDNA 3.1-Flag-Uncv 和pcDNA 3.1-Flag-vector 用Lipofectamine 2000 试剂转染HeLa 细胞。具体操作依照Lipofectamine 2000 试剂盒附带操作规程进行。转染48 h 后,用G418 对细胞进行筛选。14 d 后可见有抗性克隆细胞,在高倍镜下用枪头挑取单个细胞团种于96 孔板,继续用G418 筛选,将单克隆细胞扩大培养后用Western blot 方法检测UNCV 蛋白质的表达。
1.2.2 Western blot 检测UNCV 蛋白质的表达
收集稳定过表达UNCV 蛋白质的HeLa 细胞株,用1 × SDS 上样缓冲液裂解,沸水煮沸15 min。10% SDS-PAGE 电泳后转PVDF 膜,用含5%脱脂奶粉的TBST 溶液对PVDF 膜封闭,然后在4℃孵育一抗过夜,一抗分别为anti-Flag 单抗(1∶5000 稀释)和内参Beta-actin 单抗(1∶5000 稀释)。二抗为HRP标记的羊抗小鼠IgG (1∶5000 稀释),室温孵育1 h。Western 化学发光液孵育PVDF 膜1 min,在暗室用X 光胶片显影并定影。
1.2.3 过表达UNCV 蛋白质对于血清饥饿诱导的HeLa 细胞凋亡的影响
选取状态良好的稳定克隆HeLa 细胞,在37℃,5% CO2,饱和湿度条件下培养,待细胞密度达到100%后,洗细胞数次,用无血清的培养基培养,进行如下实验:
(1)分别在饥饿后0、54、66、78、90、102 h 收取细胞,用台盼蓝染色后进行活细胞计数。活细胞不着色,死细胞着色。每个时间点有三个重复,计数结果取平均值。
(2)分别在饥饿后0 h 和54 h 收取细胞,按照Annexin V-FITC 凋亡检测试剂盒说明书操作,用流式细胞仪检测两种稳定克隆细胞凋亡情况。
1.2.4 过表达UNCV 蛋白质对于阿霉素诱导的HeLa 细胞凋亡的影响
选取状态良好的稳定克隆HeLa 细胞,在37℃,5% CO2,饱和温度条件下进行培养,待细胞密度达到100%后,用含DOX 的完全培养液进行培养,分别在诱导后0、36、48、60、72、84 h 收取细胞,用台盼蓝染色后进行活细胞计数。每个时间点有三个重复,计数结果取平均值。
2 结果
2.1 稳定过表达UNCV 蛋白质HeLa 细胞株的鉴定
成功获得稳定过表达UNCV 蛋白质以及空载体的HeLa 细胞株,用Western blot 印迹检测稳定克隆细胞的表达(图1)。结果表明,两种稳定克隆的细胞的内参表达量基本一致,而且用anti-Flag 一抗检测后发现过表达UNCV 组细胞能够表达UNCV蛋白质,与预期结果一致。
2.2 过表达UNCV 蛋白质对血清饥饿诱导的He-La 细胞凋亡的影响
血清饥饿0 ~102 h,分别进行细胞计数。随着饥饿时间的增加,相对空载体组,过表达UNCV 组凋亡细胞数目明显减少(图2)。流式细胞术分别检测空载体组和过表达UNCV 组血清饥饿前后细胞的凋亡情况(图3),血清饥饿前两种稳定克隆细胞凋亡数目基本一致,血清饥饿后发现空载体组凋亡细胞比例为51.36%,而过表达UNCV 组凋亡细胞数目比例为12.81%。流式细胞术结果与细胞计数结果相一致。
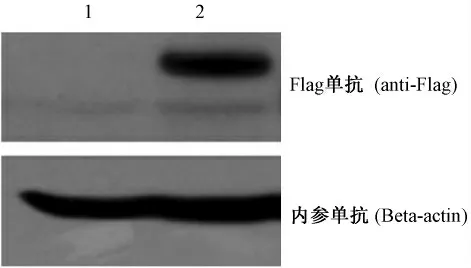
图1 稳定过表达UNCV 蛋白质HeLa 细胞株的鉴定Fig.1 Identification of the HeLa cells stably overexpressing UNCV protein

图2 细胞计数检测过表达UNCV 蛋白质对血清饥饿诱导的HeLa 细胞凋亡的影响Fig.2 Cell counting analysis of the effects of overexpressed UNCV protein on apoptosis induced by serum starvation in HeLa cells
2.3 过表达UNCV 蛋白质对阿霉素诱导的HeLa细胞凋亡的影响
DOX 为常见的化疗药物,能引起肿瘤细胞凋亡,从而起到抗肿瘤的作用[6]。稳定克隆细胞经过DOX 处理后,空载体组和过表达UNCV 组的存活HeLa 细胞数目没有明显变化(图4)。结果说明过表达UNCV 蛋白质没有影响DOX 诱导的HeLa 细胞凋亡。
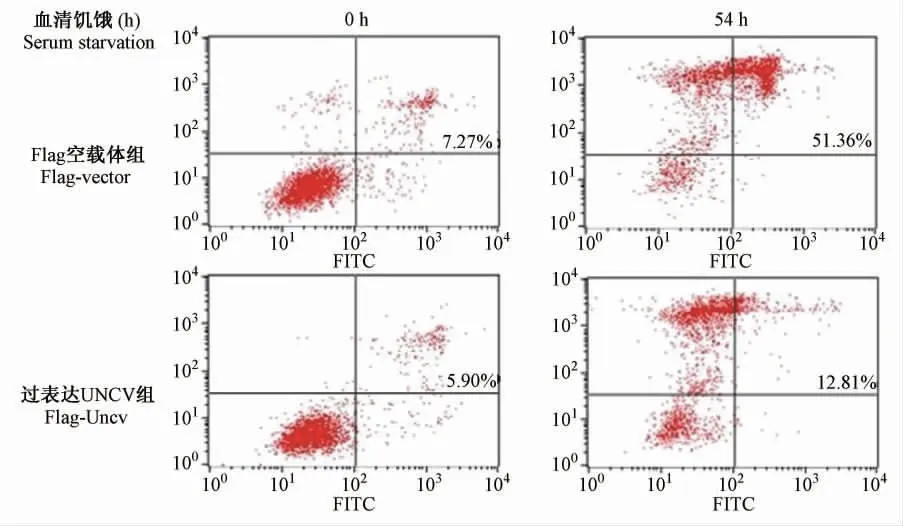
图3 流式细胞术检测过表达UNCV 蛋白质对血清饥饿诱导的HeLa 细胞凋亡的影响Fig.3 Flow cytometry analysis of the effects of overexpressed UNCV protein on apoptosis induced by serum starvation in HeLa cells
3 讨论
哺乳动物细胞主要有两种凋亡途径,一是经死亡受体介导的细胞外凋亡途径,细胞表面的死亡受体Fas 与死亡配体(FasL)结合会引起受体多聚化,从而启动细胞凋亡。FADD(Fas 偶联死亡区域蛋白Fas-associated death domain protein)与Fas 死亡结构域结合后变构,激活Caspase-8 后启动下游的Caspase 级联反应,导致细胞凋亡。第二种途径是由线粒体介导的细胞内凋亡途径。当细胞处于DNA损伤或营养供缺乏时,会激活BH3 亚族蛋白质,然后作用于Bcl-2 家族蛋白质,使Bax 和Bak 在线粒体外膜上聚集,导致线粒体外膜的通透性增加,释放出细胞色素C 和Smac/DIABLO 从而介导caspase 的级联反应,导致细胞凋亡[7]。血清饥饿和DOX 诱导的细胞凋亡都是主要通过线粒体途径,但DOX 能够直接嵌入到DNA 中形成复合物从而导致DNA 损伤,比血清饥饿引起的损伤更为有效和迅速。
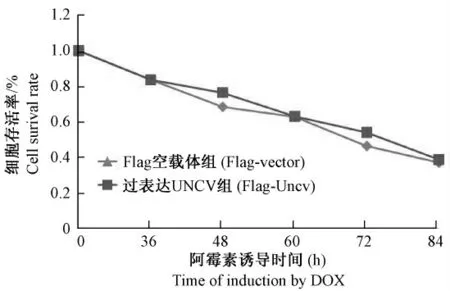
图4 细胞计数法检测过表达UNCV 蛋白质对血清饥饿诱导的HeLa 细胞凋亡的影响Fig.4 Cell counting analysis of the effects of overexpressed UNCV protein on apoptosis in HeLa cells induced by doxorubicin
一般情况下,细胞的增殖和凋亡受到严格的控制,并保持在一个特殊的平衡状态。肿瘤的发生一般都由细胞的过度增殖和凋亡异常引起,所以可以通过增加肿瘤细胞的凋亡达到治疗肿瘤的目的。目前的研究中,人类已经发现了很多与肿瘤细胞凋亡相关的基因,如p53 抑癌基因可以促进细胞凋亡[8]。在对Uncv 基因的研究中,我们发现它也能够影响与肿瘤相关的信号通路,所以我们采取过表达UNCV 蛋白质的方法观察其是否与肿瘤细胞凋亡有关。
外源DNA 导入哺乳动物细胞的技术称为细胞转染,是研究基因功能的一种重要方法,细胞转染技术分为瞬时转染和稳定转染。前者是指外源基因进入受体细胞后不整合入染色体中,但不能被复制,因此表达时间有限,通常仅一两天。后者是在前者的基础上,外源DNA 整合到受体细胞染色体上,从而可以进行复制,其子代细胞也能表达外源基因。通常会将选择性标记基因和目的基因融合表达,通过使用选择性培养基,就可以消除瞬时转染的细胞,获得目的基因成功整合到细胞染色体中的稳定克隆细胞。在本研究中,由于在血清饥饿和DOX 诱导后的观察时间较长,所以必须要采用稳定转染和G418筛选的方法获得能够稳定过表达UNCV 蛋白质的HeLa 细胞,这种细胞更利于获得可靠的实验结果。
本研究采用脂质体转染和G418 筛选的方法构建了稳定过表达UNCV 蛋白质的HeLa 细胞株。体外细胞实验证实了过表达UNCV 蛋白质能抑制血清饥饿诱导的HeLa 细胞凋亡。同时过表达UNCV蛋白质对化疗药DOX 诱导的HeLa 细胞凋亡没有明显差异。结果提示可能这是由于过表达UNCV蛋白质后促进了一些细胞生长因子的表达或分泌,从而提高了HeLa 细胞对血清饥饿诱导的细胞凋亡的抵抗能力。而DOX 诱导细胞凋亡的直接作用于双链DNA[9],相对生长因子的剥夺来说,位置处于下游,因此UNCV 没有影响DOX 诱导的HeLa 细胞凋亡。本研究结果表明Uncv 可能是治疗肿瘤的一个重要靶点。
[1]李善如,仇志华,王冬平.一种新的被毛突变小鼠及其生物学特性的初步研究[J].军事医学科学院院刊,1995,19(1):26 -28.
[2]李善如,王冬平,史燕燕,等.一种新的被毛突变小鼠遗传特性研究[J].中国兽医学报,1995,15(3):287 -290.
[3]李善如,王冬平,兰宏,等.一种新的小鼠被毛突变基因定位[J].科学通报,1999,44(7):750 -755.
[4]徐嫄,刘冰,李文龙,等.小鼠Uncv 基因克隆及真核表达[J].中国实验动物学报,2012,20(1):81 -83
[5]Ouyang L,Shi Z,Zhao S,et al.Programmed cell death pathways in cancer:a review of apoptosis,autophagy and programmed necrosis[J].Cell Prolif,2012,45(6):487 -498
[6]Wang S,Konorev EA,Kotamraju S,et al.Doxorubicin induces apoptosis in normal and tumor cells via distinctly different mechanisms.intermediacy of H2O2-and p53-dependent pathways[J].J Biol Chem,2004,279(24):25535 -25543.
[7]Taylor RC,Cullen SP,Martin SJ,et al.Apoptosis:controlled demolition at the cellular level [J].Nat Rev Mol Cell Biol,2008,9(3):231 -241.
[8]Kracikova M,Akiri G,George A,et al.A threshold mechanism mediates p53 cell fate decision between growth arrest and apoptosis.Cell Death Differ,2013,20(4):576 -588.
[9]Maruyama S,Shibata R,Ohashi K,et al.Adiponectin ameliorates doxorubicin-induced cardiotoxicity through Akt protein-dependent mechanism[J].J Biol Chem,2011,286(37):32790-32800.
