10个桉树无性系抗寒锻炼特性的研究
2013-01-14赵娟娟吴承祯
洪 伟 赵娟娟 林 晗 吴承祯*,3
(1.福建农林大学 桉树研究中心,福建 福州 350002;2.中国科学院 城市环境研究所,福建 厦门 361021;3.武夷学院,福建 武夷山 354300)
抗寒锻炼即冷适应(cold acclimation),是植物在低温作用下,抗寒力提高的过程,实质上是植物感受低温信号、调节基因的表达和代谢的环境适应过程。当气温回暖时,由抗寒锻炼获得的抗寒力会重新丧失,这个过程称为解除锻炼或脱锻炼(cold-dehardening)。植物抗寒锻炼的分子机理可以概括为低温诱导基因的表达调控,调控的水平分别为转录前、转录和转录后,其中转录前调控包括对低温等环境信号的感受和传导[1]。抗寒锻炼能诱导植物抗冻基因的表达使植物的抗冻性增强,因此对抗冻性的研究离不开对抗寒锻炼的研究。不同品种或变种,其抗寒锻炼诱导抗冻性的能力不同。不同桉树树种也具有不同的抗寒锻炼能力,Dugald M.Paton[2]对桉树的研究显示,5种桉树对0.5℃锻炼温度的反应明显不同。
电导法被普遍用于植物抗冻性的研究和抗寒品种的筛选[3]。电导法作为比较基因型间抗冻性差异的细胞生态学指标是可行的[3~6],国内外有大量关于桉树及其他植物的抗冻性研究都表明,电解质渗透率作为细胞膜伤害程度的指标,与冻害水平间具有很好的相关性;电解质渗透率配合Logistic方程对桉树的冻害发展过程作出冻害曲线,拟合出的LT50能很好的反映植物的冻害存活率[4]。Raymond等[5]对 E.regnans一个全同胞家系用电导法识别出一个抗霜冻的非加性遗传组分。Byrne M.等[6]用电导法测定亮果桉抗冻性,结合其他QTL分析方法,定位了抗霜冻的2个QTL位点。我国引种桉树(eucalyptus)尽管有上百年的历史,但不同桉树无性系的抗寒锻炼特性研究未见报道,尤其以冷库为平台探讨抗寒锻炼、脱锻炼及苗龄处理对桉树膜伤害影响国内外鲜见报道。本研究以10个桉树无性系为对象,以大型冷库为平台,通过电导率评价抗寒锻炼、脱锻炼对桉树无性系的影响,对区别桉树基础抗冻能力与冷锻炼能力、评价桉树抗冻性具有重要理论价值。
1 材料与方法
1.1 实验材料
选择10个桉树无性系作为实验材料,依次编码为XL1~XL10。实验材料为组培苗,每个无性系30株,经2个月的培育,苗高及叶量达到要求后,于2005年9~10月进行抗寒锻炼、脱锻炼及冻害胁迫处理。
1.2 实验设计
1.2.1 抗寒锻炼与脱锻炼处理法
每个无性系取生长一致的幼苗各18株。其中CK0处理和CK对照各3株,培育于常温下的苗圃中。其余植株作为处理株进入冷库进行抗寒锻炼与脱锻炼的实验。每个处理设3个重复。
整个处理过程以抗寒锻炼情况为准分为6种处理。处理1:实验开始前常温下正常生长的植株,记为CK0处理;处理2:实验全部结束后的对照株(实验日期共 32.5d),记为 CK 对照;处理 3:6℃恒温处理 10d后的处理株,记为 CH1(cold-hardening)处理;处理 4:处理3的基础上再经3℃恒温处理10d的处理株,记为CH2处理;处理5:处理4的基础上再经0℃恒温处理10d的处理株,记为CH3处理;处理6:处理5的基础上再经脱锻炼处理的处理株,记为DH(De-hardening)处理。脱锻炼处理为5℃~25℃逐渐升温处理,即5℃、10℃、15℃、20℃、25℃依次各 12h的处理。
1.2.2 冻害胁迫处理方法
锻炼和脱锻炼后进行冻害胁迫,冻害胁迫时间为当天 8:00~16:00,实验株转入冷库后于 8:00、10:00、12:00、14:00 和 16:00 时分别将温度调为-2℃、-4℃、-6℃、-8℃、关机;降温速度为 5℃/min,每个温度处停留2h,于16:00时胁迫处理完毕。每阶段低温胁迫处理结束后,分别对处理株采集叶片并测定电解质渗透率。
以上锻炼、脱锻炼及冻害处理实验均在福建农林大学福建省农副产品保鲜技术工程中心自制的冷库(共1~8号库)内进行。该冷库的实验条件为,光照时间每日8:00~17:00,光照强度150W/m2,光源为白炽灯;水分条件:土壤保持湿润,空气湿度为80~100%。实验期间不进行施肥或施药处理。
1.3 半致死温度的测定
1.3.1 电导率测定
零下低温胁迫处理中,于当天的8:00、10:00、12:00、14:00及16:00共5个整点时间分别对处理株顺序采集+3~+7叶位的叶片,作为零下低温胁迫各阶段的处理样品测定其电解质渗透率的变化情况[3~6]。样品经充分冲洗后再经单蒸水润洗,然后用1cm直径的打孔器分别从每份样品叶中随机打取10个小圆叶片 (避开叶脉及叶缘),放入10ml离心管中,加双蒸水8ml,真空渗入3h,每隔1h取出离心管振荡一次。待叶片下沉后,静置10min,测定处理电导值B;后置沸水浴中10min,冷却后测煮后电导值C。并依下式计算电解质渗透率[3~6]:

1.3.2 半致死温度计算
根据电解质渗透率进行回归分析,得到logistic方程(y=k/(1+ea-rt)),求得 y=k/2 时的 t值(温度)作为桉树冻害胁迫的半致死温度[3~6]。采用电解质渗漏达到50%的低温点作为植物的半致死温度,即在“温度-相对电导率”的S型曲线上表现为曲线的拐点所对应的温度,以直观、准确地反映植物的抗寒能力[7]。
1.4 数据处理
数据分析均采用Spss10.0与Excel 2000处理完成,Logistic曲线的拟合和拐点温度的求解由自编的Qbasic程序计算。
2 结果与分析
2.1 抗寒锻炼处理半致死温度的差异显著性分析
分析测定10个无性系各6种处理的电解质渗透率,获得半致死温度及曲线拟和R2值(表1)。对不同桉树无性系半致死温度进行方差分析,表明6个处理间的差异性达到极显著水平(p<0.05),但是不同无性系间未达到显著水平(p>0.05)。表明,半致死温度可用于比较6种不同处理的抗冻性差异,但不适于区别不同桉树无性系间抗冻性的差异。尽管不同树种间的抗冻性差异一般较明显,但树种不同无性系间的抗冻性差异比树种间的差异小得多。
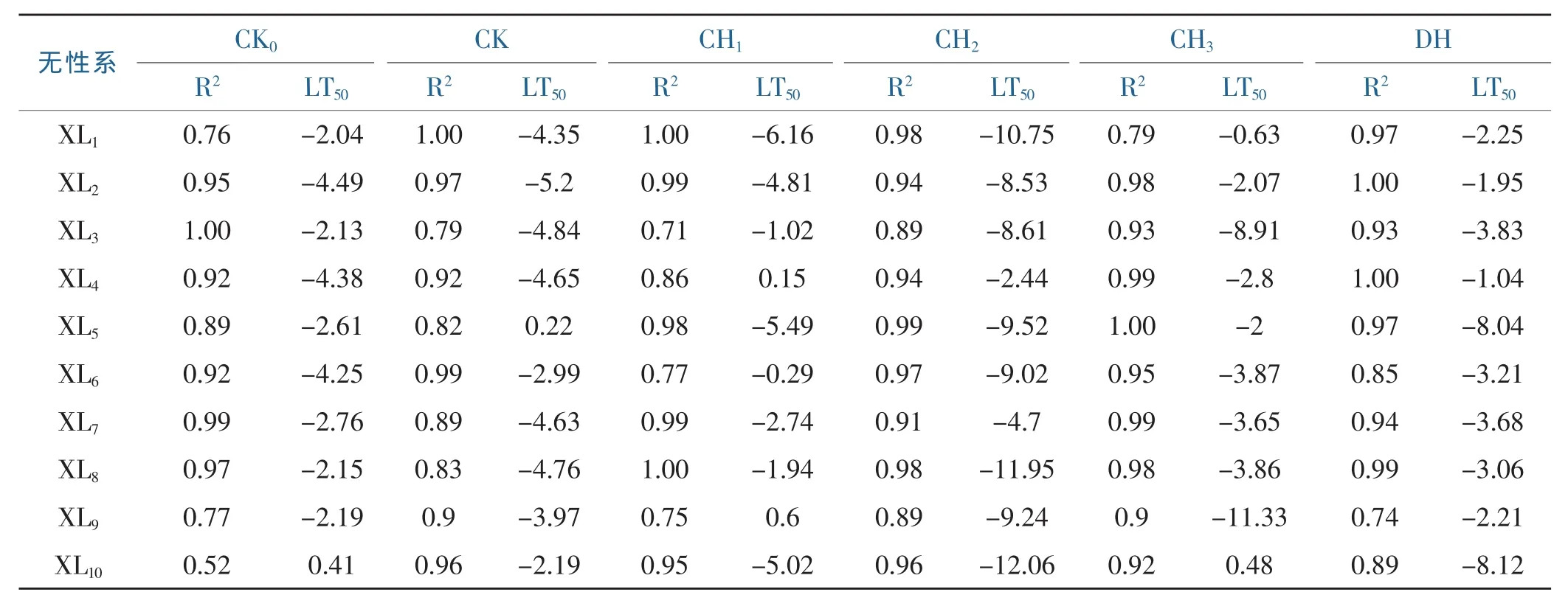
表1 10个桉树无性系的半致死温度(LT50,单位℃)Table 1 The temperatures for 50%survival rate of eucalypt clones(LT50,unit℃)
对10个桉树无性系6个处理的半致死温度进行配对T检验(表2),表明3种锻炼处理中只有CH2锻炼处理与CK对照处理的差异性达到极显著水平,而CH1、CH3锻炼处理与CK对照处理的差异均不显著,即3个锻炼处理水平中以CH2处理增强桉树无性系抗冻性的效果最好。但DH脱锻炼处理与CK对照处理的差异不显著,因此CH2锻炼处理所获得的抗冻性经脱锻炼处理后将消失。3个锻炼处理中,CH2与CH1、CH32个锻炼处理之间的差异性均达到极显著或显著水平,但CH1、CH3之间的差异性不显著,进一步说明CH2抗寒锻炼处理明显比CH1、CH32种抗寒锻炼处理更能提高桉树的抗冻性。
2.2 抗寒锻炼对桉树无性系抗冻性的影响
研究表明抗寒锻炼可以增强大部分桉树无性系的抗冻性,但是不同无性系对相同抗寒锻炼处理的反映明显不同(图1),这与Dugald M.P.[2]的研究规律相一致。DH处理使大部分无性系的抗冻性重新降低,但是XL5和XL10的半致死温度非但没有上升反而下降,可见这2个无性系在0℃胁迫下抗冻性上升 (图1),因此该脱锻炼处理虽然使大部分无性系达到脱锻炼的效果,但是还有部分无性系未脱锻炼。

表2 不同低温处理间半致死温度的配对T检验结果Table 2 Paired-samples T test results of LT50 in different chilling treatments
总体而言,LT50反映的不同低温处理对桉树无性系的抗冻性影响规律较为复杂,CH2锻炼处理使大部分无性系的抗冻性大幅上升 (如 XL1、XL2、XL5、XL6、XL8、XL10),但也使一些无性系的抗冻性降低 (如XL4)。CH1和CH3锻炼处理对大部分桉树无性系的抗冻性没有正面的影响,而是使其抗冻性大幅降低,或者不影响其抗冻性。苗龄相差1个月的苗龄处理,比苗龄更大的那组对照株抗冻性降低。

Li[8]将马铃薯抗冻基因型分为RA(冷抗可锻炼型)、RUA (冷抗不可锻炼型)、SA (冷敏可锻炼型)、SUA(冷敏不可锻炼型)4种类型,并推测植物的基础抗冻性与锻炼能力具有不同的遗传机理。郭玉华等[9]的研究也支持此观点,其用存活率和离子渗透率检测法测定了18个小麦品种,结果表明,离子渗透率检测分辨率最高的温度是-10℃,抗寒性强的品种有不同的抗寒机制,基础抗冻能力与冷锻炼能力对最终抗冻性的贡献有很大差别。本文的实验结果支持相同的结论和推测。
2.3 桉树无性系处理间半致死温度差值的变化规律
将锻炼、脱锻炼、未锻炼3种不同处理的半致死温度相减,以比较处理间的差异(表3)。以所得差值作为不同处理桉树无性系间抗冻性的差异水平,对不同处理间的抗冻性水平进行比较分析。其中“CK-CK0”是常温下正常生长株在实验结束后1d(CK)与实验前1d(CK0)2个时间点间的半致死温度差值,代表常温下正常生长株经过1个月的生长之后半致死温度的变化情况。由于CH2锻炼处理、CH3锻炼处理与DH脱锻炼处理的处理时间分别为20d、30d和32.5d,所以其生长引起的抗寒性与CK相似,选用CK对照与CH2、CH3和 DH 进行比较,“CH2-CK ”、“CH3-CH2”和“DHCK”分别代表二者间的半致死温度差值,是剔除生长因素的影响后CH2、CH3锻炼处理和DH脱锻炼处理对桉树半致死温度的影响情况。同理,由于CH1锻炼处理为10d,所以其生长引起的抗寒性与CK0相似,借用与CK一样未做低温处理的CK0处理(苗龄处理)与CH1进行比较。“CH1-CK0”即二者间的半致死温度差值,代表剔除生长因素的影响后CH1锻炼处理对半致死温度的影响情况。
由于10个无性系的对照株在实验期前、后的半致死温度差值全部为负值,可知半致死温度即抗冻性随着苗龄的增加而上升(表3)。CH2锻炼处理使无性系的抗冻性明显增强,平均使各无性系的半致死温度降低了4.95℃。CH1和CH3锻炼处理可以增强某些无性系的抗冻性,但是效果不明显。CH1锻炼处理使无性系的半致死温度平均下降了0.013℃,CH3处理使半致死温度平均下降了0.128℃。进一步证明,大部分无性系以CH2抗寒锻炼处理(即6℃10d+3℃10d)增强抗冻性的效果最佳(表3)。

表3 不同锻炼处理与对照处理的半致死温度差值表Table 3 Deviation values between chilling treatment and CK
2.4 桉树无性系抗寒锻炼特性与抗冻性机理分析
根据郭玉华等[9]的方法,这里也将抗寒锻炼前与抗寒锻炼后桉树无性系的半致死温度进行排序并分别聚为3类。应用SPSS10.0对抗寒锻炼前或未锻炼(CK0)10个桉树无性系的半致死温度值(表 4),进行K-均值聚类分析,其中1类的聚类中心为-4.37、2类的聚类中心为-2.31、3类的聚类中心为0.41(表4)。经计算抗寒锻炼处理后各无性系抗冻性的最大增强值即CH1-CK0、CH2-CK和CH3-CK中的最小值 (即LT50降低的最大值),可进行增强值大小的排序和聚类分析(表4),其中1类的聚类中心为-8.54、2类的聚类中心为-4.96、3类的聚类中心为0.90。
基于冷抗与冷敏性,比较各无性系在锻炼前的半致死温度及抗寒锻炼引起的抗冻性增加值 (表4)和CH3锻炼引起的抗冻性增强值(表3),表明锻炼前抗冻性聚类(CK0)为“1”的作为10个无性系中的冷抗无性系,聚类为“3”的作为冷敏无性系;锻炼后ΔLT50聚类为“1”的是锻炼性强的无性系,为“3”的是锻炼性弱的无性系;聚类为“2”的均为中间型的无性系。
根据锻炼后的maxΔLT50与锻炼前的CK0对不同无性系的抗冻性机理进行分析,XL4的CK0为“1”类,而其maxΔLT50为“3”类,所以XL4属于冷抗不可锻炼无性系;XL7的 CK0为“2”类,而其 maxΔLT50为“3”类,所以是中性抗冷不可锻炼的无性系;XL5、XL8、XL9和XL10均为可锻炼的无性系,maxΔLT50都为 “1”类,除XL10为冷敏可锻炼无性系外其他无性系为中性抗冻可锻炼无性系;其它4个无性系均为中等可锻炼的无性系,其中XL1、XL3为中性抗冻中等可锻炼无性系,而XL2和XL6为冷抗可锻炼无性系。

表4 CK0处理桉树无性系LT50与抗寒锻炼后桉树无性系maxΔLT50的聚类分析Table 4 Results of cluster analyses for the maxΔLT50 after chilling treatments functioning as cold-hardening processes and LT50 of SA treatment of eucalypt clones
3 结论与讨论
研究10个桉树无性系苗龄相差1个月的2种未锻炼植株在冻害胁迫后半致死温度的变化情况,发现所有无性系的抗冻性均在1个月后有所增强,半致死温度平均降低了1.08℃,以苗龄更大的那组未锻炼处理的抗冻性较高。而且不同的桉树无性系根据半致死温度进行抗冻性排序的结果,在1个月的生长期后有较大的变化。可见桉树不同无性系的抗冻性会随着苗龄的增长而变化,并且变化的幅度和抗冻性排序的结果都有变化。本实验选用苗龄相同的无性系进行抗寒锻炼性研究,并设置了1个苗龄处理比较生长与抗寒锻炼对抗冻性的影响,在抗寒锻炼的分析过程中剔除生长对抗冻性的影响进行分析,使抗冻性评价和抗寒锻炼性分析的准确性大大提高。
半致死温度计算结果表明,6个处理之间的差异性达到极显著水平,但是不同无性系间半致死温度的差异未达到显著水平,所以半致死温度用以比较不同处理的抗冻性差异是适宜的。不同低温处理对桉树无性系的抗冻性影响规律较为复杂。大多数的桉树无性系在6℃10d+3℃10d的CH2锻炼处理后,抗冻性比未锻炼的对照株有大幅度增强,半致死温度平均降低了4.95℃;但是也有一些无性系的抗冻性降低。CH1和CH3锻炼处理对大部分桉树无性系的抗冻性没有正面的影响,而是使其抗冻性大幅降低,或者不影响其抗冻性。脱锻炼处理对桉树无性系的抗冻性恢复能力也有不同。根据锻炼后的maxΔLT50与锻炼前的CK0对桉树不同无性系的抗冻性,借鉴Li[8]对抗冻性基因型划分的4种类型,认为XL4属于冷抗不可锻炼无性系、XL7属冷敏不可锻炼的无性系,XL2和XL6为冷抗可锻炼无性系;其它6个无性系为冷敏可锻炼无性系( 其中 XL5、XL8、XL9属中性抗冻可锻炼无性系,XL10属冷敏可锻炼无性系,XL1、XL3属中性抗冻中等可锻炼无性系)。
植物抗冻性与其抗寒锻炼性有很大的关系,这种抗寒锻炼程度在自然界表现为零下低温冻害前零上低温的温度和持续时间,因此不同地区不同气候下形成的抗寒锻炼条件是不同的。Dugald M.P.[2]研究表明,热带种棉芝老桉(Eucalyptus.deglupta)不能响应低温锻炼,不管它锻炼时间多长,在-8℃冰冻下所有植株都死亡;而多枝桉 (Eucalyptus.viminalis)和稀花桉(Eucalyptus.Pouciflora)经 4~5d锻炼后有半数以上的植株足以忍耐-8℃冰冻并且无伤害,这2种桉树的最大抗冻力分别产生于锻炼后10d和20d,且稀花桉的抗性比多枝桉大得多;其余几种桉树的锻炼启始速率及最终达到的抗冻水平介于稀花桉和棉枝老桉之间。因此,树种抗寒性锻炼,其抗冻性差异较大,本研究也表明抗寒性处理不同桉树抗冻性排序不一样。同理,桉树抗寒锻炼受体生长期不一样,其抗冻性排序也将变化。因此桉树抗寒无性系的筛选应该选用什么实验材料、如何进行抗寒性实验才能较好地反映成年桉树无性系的抗冻性还有待进一步探讨。
[1]Jermstad K.D.,Bassoni D.L.,Wheeler N.C.,et al.Mapping of quantitative trait loci controlling adaptive traits in coastal Douglas-fir.II.Spring and fall cold-hardiness[J].Theor Appl Genet,2001,102:1152-1158.
[2]Dugald M.P.Is there participation of growth regulating substance in cold-resistance of eucalypts 见:刘祖祺,王 洪春.植物耐寒性及防寒技术[M].北京:学术书刊出版社,1990,65-70(in Chinese).
[3] 廖国华,陈俊河,李鹏程等.电导法在桉树抗寒评价中的应用[J].林业科技开发,2002,16(4):17-19.
[4]Rajashekevar C,Gusta L V,Burke M J.Forest damage in hardy herbaceous species [M].New York:Academic Press,1979,255-274.
[5]Raymond CA.Harwood CE,Owen JV.A conductivity method for screening population of eucalyptus for frost damage and frost tolerance[J].Aust JBot,1986,34:377-393.
[6] Byrne M,Murrell JC,Owen JV,Kriedemann P,Williams ER.Moran GF Identification and mode of action of quantitative trait loci affecting seedling height and leaf area in Eucalyptus intense[J].Theor Appl Genet,1997,94:674-681.
[7] 朱根海.应用方程确定植物组织低温半致死温度的研究[J].南京农业大学学报,1986,9(3):11-16.
[8] Li P H,Palta J P,Chen H H.Freezing stress in potato.In:Low temperature stress in crop plant[M].New York:Academic Press,1979,291-303.
[9] 郭玉华,H.Wisniewska,J.Chelkowski.不同小麦基因型抗寒性与冷适应特征的评价 [J].沈阳农业大学学报,2000,31(6):541-545.
