油茶芽苗砧嫁接体愈合过程AFLP分析
2012-12-28冯金玲杨志坚
冯金玲,杨志坚,陈 辉
油茶芽苗砧嫁接体愈合过程AFLP分析
冯金玲,杨志坚,陈 辉
(福建农林大学, 福建 福州350002)
为了探讨油茶芽苗砧嫁接体愈合过程中嫁接口和接穗是否发生基因变化,利用AFLP对不同发育阶段的油茶芽苗砧嫁接口,油茶芽苗砧愈伤口以及接穗母树、砧木和接穗萌发芽的基因进行分析。结果表明:油茶芽苗砧嫁接口发育过程中,条带数变化相对比较大,特别是在嫁接移栽后的第 4、16、24、35 天,愈伤口条带数的变化是在移栽后第 0 天至第 18 天间;接穗母树、接穗萌发芽和砧木存在 13 条特异带,10 条在接穗萌发芽DNA 中存在,其中 5 条为接穗萌发芽特有条带,5 条在砧木茎中存在。
油茶;芽苗砧嫁接;基因;AFLP
油茶Camellia oleifera 是我国重要的木本油料树种,已有 2 300 多年的栽培历史[1]。大力发展油茶生产,不仅是生态环境建设的需要,而且是不断满足人们生活所需和调整农业产业结构、发展农村经济、增加农民收入的重要途径[2]。油茶芽苗砧嫁接具有操作简单、繁殖系数大、结实早、能保持母树优良性状等优点,在油茶生产中是良种繁育的主要途径[3]。嫁接是否引起植物基因变化目前存在两种看法,一种认为嫁接不会引起植物基因的变化。如周瑞金等人运用携带外源基因nptⅡ的苹果品种接体和未进行基因转化的苹果组培苗作砧木, 通过试管微嫁接,检测结果表明外源nptⅡ基因只在转化植株体内表达, 而不通过嫁接在接穗和砧木间进行传导[4]。另一种观点认为嫁接能引起植物变异。如Hirata等人在辣椒嫁接实验中认为,嫁接能诱导辣椒嫁接植株性状产生变异,包括果形、株型以及辣椒素含量等,并能稳定遗传27代[5-7]。迄今为止,尚未见到芽苗砧嫁接是否引起嫁接体基因变化的研究报道。为此,本实验运用AFLP方法[8-10],分析油茶芽苗砧嫁接体愈合过程中接穗和砧木是否发生基因变化,旨在判断芽苗砧嫁接能否保持接穗的优良品质和探索一种快捷育种方法。
1 材料与方法
1.1 材 料
2009 年6月2日于田间实验室进行油茶芽苗砧嫁接。砧木的种子来源于福建省桐口林场的优良无性系,砧木茎段截留1.5 cm,根部留6 cm左右;接穗取自同一株优树上健壮的半木质化的当年生枝条,长2 ~ 3 cm,一芽一叶。取10g左右的接穗茎和砧木茎,保存于 -80 ℃ 冰箱。对照实生苗与砧木进行相同的截茎断根处理。处理后立即栽植于预先整好的苗床。株行距3 cm× 10 cm。栽植时压实土壤,籽粒露在畦面,浇透水,盖上白色透明塑料薄膜。棚内的温度保持在28 ~ 30 ℃。
6月2日实验处理后移栽至第26 天每 2 d取1次样,共取14次;至第35 天每隔3 d取1次样,共3次;至第45 天每隔5 d取1次样,共2次;至第55 天隔10 d取1次,共1次,合计取20次样,每次都采用完全随机方法进行取样。取样后植株在流水中清洗干净,截取油茶芽苗砧嫁接苗的嫁接口和实生苗的愈伤口1 ~ 1.5 cm 的茎段以及萌发的接穗芽(见表1),保存于 -80 ℃ 冰箱。
1.2 DNA提取
每个处理随机抽取 15 株,约1.5g的材料,用CTAB 法提取总 DNA[11-12],上海生工的DNA纯化试剂盒纯化 DNA,测定DNA 浓度,并用1%Agarose胶检测 DNA 的质量。稀释至 20 ng·μL-1,-20℃保存备用。
1.3 AFLP分析
设计选取 8 对引物组合(见表2)进行检测。EcoR I 和 Mse I 两种限制性内切酶、EcoRI和MseI 接头、引物、T4-DNA 连接酶、Taq DNA 聚合酶及 DNA ladder marker 均购自纽英伦生物技术(北京)有限公司。

表1 参试材料编号†Table 1 Numbers of the materials in experiment

表2 油茶AFLP 引物序列Table 2 AFLP primers sequences of Camellia oleifera
1.3.1 酶 切
选取 EcoR I/Mse I 酶切组合。酶切反应体系 20 μL,EcoRI 为1 U,MseI为3 U,DNA 为150 ng,混匀后 37 ℃ 水浴 8 h,酶切后,80 ℃ 水浴 10 min 灭活。
1.3.2 连 接
反 应 体 系 20 μL,DNA 酶 切 液 10 μL,EcoRI 接头和 MseI 接头各 5 pmol,T4-DNA 连接酶 7 U,10 mmol·L-1ATP 0.2 mmol/L,1 μL 5× 酶切连接缓冲液。上述混合液 16 ℃ 保温过夜,连接结束后,80 ℃ 水浴 10 min 灭活,-20 ℃ 保存备用。
1.3.3 预扩增
反应体系 20 μL,DNA 连接液 4 μL,dNTP 终浓度为 0.2 mmol·L-1,Primer E-A 为 2 pmol,Primer M-C 为 1 pmol,Mg2+终 浓 度 为 2.5 mmol·L-1,rTaq聚合酶 1.5 U。混匀后在 Eppendor 梯度 PCR仪上进行扩增。程序为 95℃ 4 min,95℃ 30 s,56℃ 30 s,72℃ 1 min,循环 30 次,最后 72℃再延伸 10 min。PCR 后吸取 10 μL 预扩增产物和 2 μL Loading buffer 混合,在 2 % 琼脂糖凝胶中检测预扩增的效果,剩余的预扩增产物置于-20 ℃ 冰箱保存。
1.3.4 选择性扩增
反应体系20 μL,取5 μL稀释60倍的预扩增产物,Primer E-ANN为8 pmol,Primer M-CNN为 3 pmol,dNTP 终浓度为 0.2 mmol·L-1,Mg2+终浓度为2.5 mmol·L-1,rTaq聚合酶1.5 U。混匀后在Eppendor 梯度 PCR 仪上进行扩增。程序为95℃ 4 min,而后按照95℃ 30 s,65℃ 30 s(每个循环降低 0.7 ℃ ),72 ℃ 1 min,循环12次,再按照95℃ 30 s,56℃ 30 s,72℃ 1 min,循环23次,最后72℃延伸10 min。PCR 后吸取 10 μL 预扩增产物和 2 μL Loading buffer 混合,在 2 % 琼脂糖凝胶中检测选择性扩增效果。
1.3.5 结果检测
选择性扩增产物与加样缓冲液等体积混合液,95 ℃ 变性 6 min,置冰上冷却,吸取 3.5 μL 混合液上样,在 85 W 下用 6% 的聚丙烯酰胺凝胶电泳至二甲苯青离底部 1~2 cm 时停止,分离扩增产物,银染检测。
1.4 数据分析
人工记录电泳条带,根据DNA电泳分子量标准(Marker)计算出各片段的大小,按扩增条带的有无分别计为1和0,形成数据进行分析处理。
2 结果与分析
2.1 AFLP引物筛选
本实验用 8 个 EcoRI 引物和 8 个 Mse I 引物组合成 64 对 AFLP 引物(见表3),以 1 号材料的DNA 为模板进行扩增。扩增产物用6 % 变性聚丙烯酰胺凝胶进行检测,结果见图 1。本实验产生的AFLP 聚丙烯酰胺凝胶图 200 bp 以下的条带发虚,因此选择分析的条带在 200~500 bp 之间。由表3可得,不同引物产生的条带数差异很大,在 15~48 条之间,其中 E5M4 组合引物产生的条带最小,15 条;而 E1M6 组合引物却产生了 48 条引物。同时不同引物组合扩增的效果也不同,因此选择条带清晰、且稳定性和多态性好的引物组合。结合扩增的条带性和效果,本实验选择 E1M1 和 E1M6 两对引物组合。

表3 AFLP-PCR 引物扩增的结果统计Table 3 The results of the primes AFLP-PCR amplification
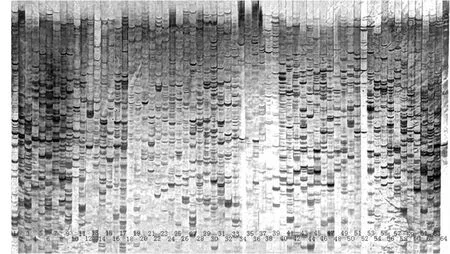
图1 引物筛选变性聚丙烯酰胺凝胶电泳Fig.1 The primers screening tested by denatured polyacrylamide gel electrophoresis
2.2 嫁接体愈合过程AFLP分析
愈合过程中的嫁接口和愈伤口AFLP选择性扩增图谱见图2。样品号 1~20 为油茶芽苗砧嫁接口发育过程的材料。由表 4 可见,在 E1M1 的引物组合中,条带数变化较大,其中条带数最少的是在样品 8和 9 中,为 43条;而在样品 13、17 和 18 中条带数最多,为 48 条;带数变化最大是在样品 3 号与样品 4 号之间,为3 条。在 E1M6的引物组合中,条带数变化较小,在 28 ~ 31 之间,呈缓慢上升后下降,其变化分别在样品7、样品9和样品17中。综合以上可见,在油茶芽苗砧嫁接愈合过程中,嫁接口的基因存在变化,而且在样品 3、样品 9、样品 13 以及样品 17 即嫁接移栽后的第 4 天 、第 16 天第 24 天及第 35 天可能是油茶芽苗砧嫁接愈合的关键时期。

图2 E1M1和E1M6 引物对愈合过程中的油茶芽苗砧嫁接口 AFLP 选择性扩增图谱Fig.2 Map of AFLP selection amplification in prime E1M1and E1M6 for development of Camellia oleifera nurse seed grafted union
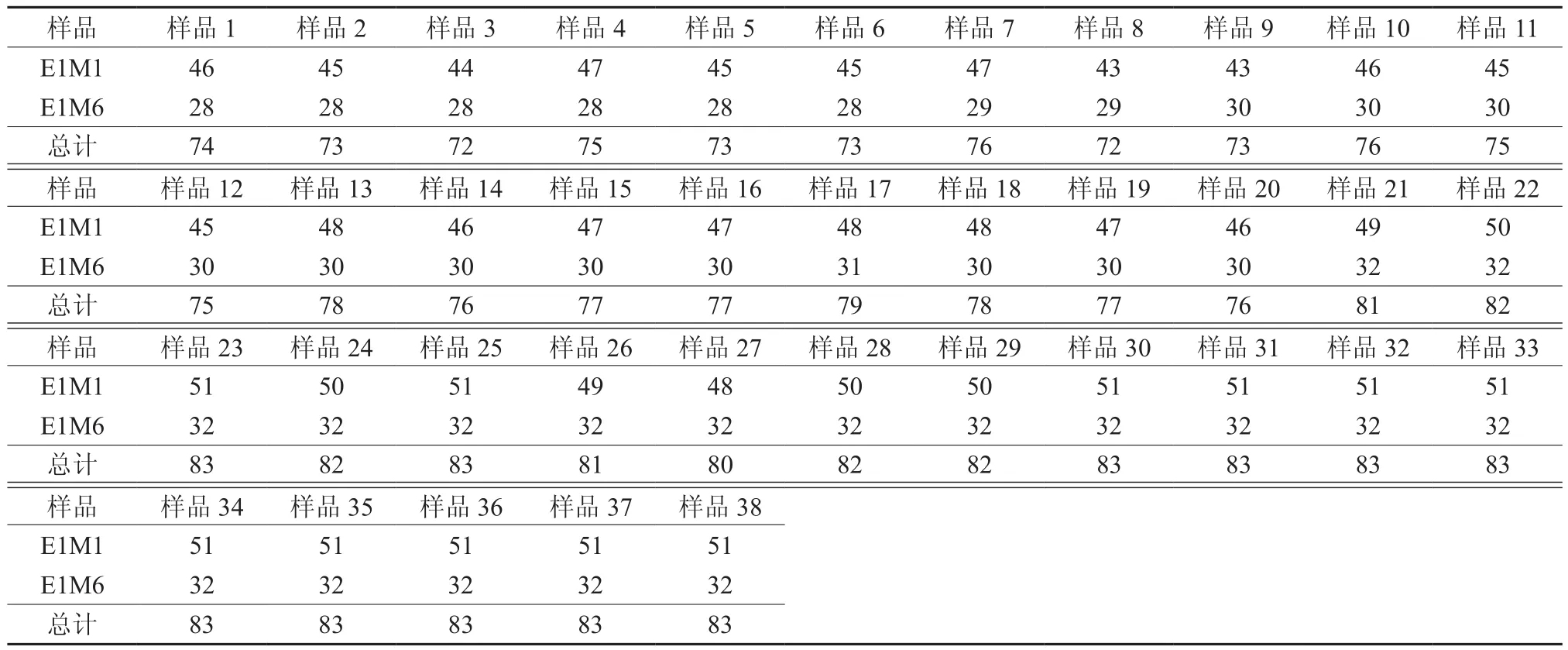
表4 引物对愈合过程中的油茶芽苗砧嫁接口扩增的条带数Table 4 The bands amplified by AFLP primers among development of Camellia oleifera nurse seed grafted union
样品号 21~38 是油茶芽苗砧愈伤处理后愈合过程的材料。由表 4 可见,虽然在 E1M1 的引物组合中存在 3 个条带数的变化,其变化是在样品 21 至样品 29 之间即移栽后第 0 天 至第 18 天间,有可能此期是油茶芽苗砧愈伤口愈合的过程。但在 E1M6 的引物组合中没有条带数的变化。
2.3 接穗萌发芽AFLP分析
样品 39、40 和41 分别代表着接穗母树茎、接穗萌发芽和砧木茎。由图3可见,在 E1M1 的引物组合中,接穗萌发芽的条带数为51,比接穗母树茎的条数带多6 条,比砧木茎条带数多2 条。在 E1M6 的引物组合中,接穗萌发芽的条带数为30,比接穗母树茎的条数带多2 条,比砧木茎条带数多1 条。总之三者条带数大小排序分别为:接穗萌发芽、接穗母树茎、砧木茎。

图3 E1M1和E1M6 引物对接穗萌发芽 AFLP 选择性扩增图谱Fig.3 Map of AFLP selection amplification in prime E1M1 and E1M6 for the buds from the scion
从表5可得在接穗母树茎、接穗萌发芽和砧木茎中总共有13条特异条带。10个特条带在接穗萌发芽的DNA中存在,接穗母茎DNA中不存在,其中有5条在砧木茎DNA中存在,剩余的5条是接穗萌发芽特有的条带。剩下的3条特异带,1条为接穗母树茎特有条带;1条为砧木茎特有条带;1条在接穗萌发芽的DNA中不存在,但在接穗母茎和砧木茎DNA中存在。
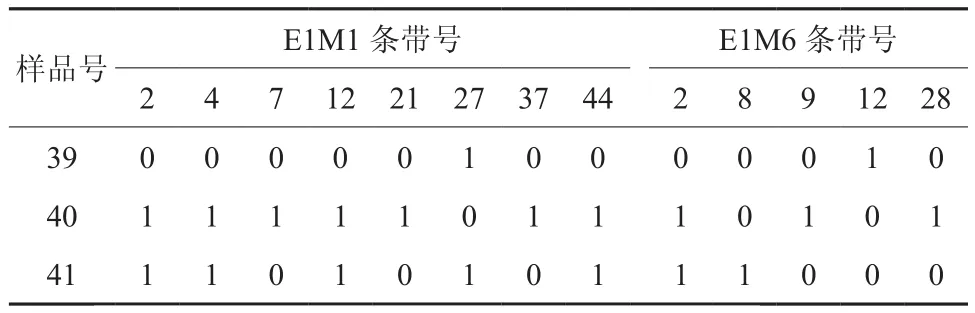
表5 2对引物在材料39、40和41中的AFLP特异带Table 5 Data of the AFLP specific bands with 2 pairs of AFLP primers among material from 39 to 41
3 讨 论
DNA 是遗传信息的载体,信息包含于 DNA的碱基排列顺序之中。DNA 分析技术在原理上可大致分为两大类,一类是直接测序,分析一些特定基因或DNA 片段的核苷酸序列,度量这些片段DNA 的变异性;另一类是检测基因组的一批识别位点,从而估测基因组的变异性[13]。AFLP就是作为一种行之有效的通过检测基因组识别位点来判断基因组变化的分子标记。一条AFLP带的“有”或“无”反映了这一位点的 DNA 序列的差异[14]。AFLP 分析中,每一条扩增带对应着基因组 DNA分子上的一个位点。样品之间带型的差异反映了DNA 水平上酶切位点分布的差异,所以可将每一扩增片段看作基因组的一个特征,而且这种特征数目不受环境条件的影响,非常稳定可靠[15]。
嫁接愈合是异种植物或同种植物的细胞、组织或器官相互影响与作用结合成一个有机整体的过程。愈合的过程一般都经历隔离层形成期、愈伤组织分化增殖期、形成层分化与连接期及输导组织分化与连接期这4个阶段[16-17]。油茶芽苗砧嫁接口在愈合过程中AFLP 的条带数发生变化,特别是在嫁接移栽后的第 4 天 、第 16 天、第 24天及第 35 天,但油茶芽苗砧愈伤口的条带数在移栽后第 0 天至第 18 天间发生变化,有可能是嫁接口和愈伤口在阶段性的发育中基因组发生了甲基化。DNA甲基化水平在愈伤中最低,再生过程上升[18]。因此在嫁接移栽后的第4 天,嫁接口的总DNA条带数最少,为72条,此阶段有可能是愈伤组织分化增殖期;而接移栽后的第35天,嫁接口的总DNA条带数最多,为79条,此阶段有可能是输导组织分化与连接期。在植物发育过程中原始DNA 序列并未改变,而是基因组DNA序列以外的因素调控特定基因表达,当基因处于表达水平时,甲基化状态一般较低,而生长发育需要关闭时,就会发生重新甲基化,抑制转录并终止其表达[19]。这正好说明了AFLP条带数的波动变化。同时DNA甲基化水平可能与形态建成相关,随着年龄增加,DNA甲基化水平呈上升趋势[20],因此无论是嫁接口还是愈伤口的愈合过程,AFLP条带数虽波动变化,但总体上升。
接穗萌发芽DNA 中11条特异条带,5条为接穗萌发芽特有条带,5条在砧木茎中存在,1条在接穗萌发芽DNA 中不存在,在接穗母树茎和砧木茎的DNA都有存在。接穗萌发芽特异条带的变化可能存在三种原因,一种原因可能是DNA发生了甲基化,从而调控在嫁接口愈合背景下接穗芽的发育;另外一种原因可能是接穗和砧木存在着基因的交流。陈红和王永清[21]在番茄自体嫁接与番茄与茄子的异种嫁接的接合部中均出现了特征带,认为嫁接接合部愈伤组织细胞间的相互作用导致了一定的遗传变异的结果类似。李明和姜世玲等[23]也认为基因可以通过嫁接面在拟南芥体内双向传递[22]。嫁接体的砧木与接穗的 DNA 发生交流其原理可能是在木质化和死亡的细胞中,DNA 穿过细胞壁和细胞间隙向维管束转移,通过维管束砧木与接穗的 DNA 发生了转移,并在植株生长点先整合。最后一种可能是接穗受到嫁接引起的环境压力的影响,活化了转座子,而引起变异[24]。
接穗萌发芽DNA特异条带的出现是由甲基化,或者嫁接压力引起芽变异,或者嫁接愈合过程中接穗和砧木基因交流引起,还是三者共同作用或其它原因引起的需作进一步研究。同时还需进一步研究接穗萌发芽DNA的特异条带,是引起什么功能基因变化,这种变化能否稳定遗传。如果可以稳定遗传又如何做选择性育种。
[1] 周素梅,王 强.我国茶籽资源的开发利用及前景分析[J].中国食物与营养,2004,(3):13-16.
[2] 袁德义,彭邵锋,邹 锋,等.油茶种子脂肪酸与游离氨基酸的分析[J].中南林业科技大学学报,2011,31(1):77- 79
[3] 范成民,董丽芬,朱 帜,等.核桃芽苗砧嫁接方法研究[J].西北林学院学报,2008, 23 (4): 109-111.
[4] 周瑞金,杜国强,师校欣.标记基因nptⅡ在转基因苹果嫁接砧穗间无相互传导[J].园艺学报,2006,33 (6):1329-1330.
[5] Yagishita N, Hirata Y, Mizukami H, et al.Genetic nature of low capsaicin content in the variant strains induced by grafting in C.arnuum L.Euphytica[J].Arnuum L.Euphytica,1990, 46:249-252.
[6] Taller J, Hirata Y, Yagishita N, et al.Graft-induced genetic changes and the inheritance of several characteristics in pepper(Capsicum annuum L.) [J].Theor.Appl.Genet.,1998, 97:705-713.
[7] Taller J,Yagishita N, Hirata Y.Graft-induced variants as a source of novel characteristics in the breeding of pepper (Capsicum annuum L.) [J].Euphytica,1999, 108:73-78.
[8] Clement D,Risterucci A M, Motamayor J C,et al.Mapping quantitative trait loci for bean traits and ovule number in Theobroma cacao L.[J].Genome,2003, 46(1):103-111.
[9] 侯文秀,李景富,刘剑峰,等.番茄成熟突变体rin 基因的AFLP 分子标记[J].东北农业大学学报,2010,41(2):20-24.
[10] 孟庆文,崔卫东,白光强,等.红景天种内遗传多样性分析AFLP 方法建立[J].新疆农业科学,2008,45 (1):88-92.
[11] 彭邵锋,张党权,陈永忠,等.14个油茶良种遗传多样性的SRAP分析[J].中南林业科技大学学报,,2011,31(1):80- 85.
[12] 周李华,赵超艺,罗军武,等.茶树种质资源AFLP分析中DNA模板快速制备方法[J].食品科学,2007,28(2):193-195.[13] 李 璇,王晓东.AFL P 的研究[J].甘肃教育学院学报(自然科学版 ),2004,18(1):58-61.
[14] 邓建云,李建强,黄宏文.一株具有特异AFLP指纹图谱的杜仲古树[J].武汉植物学研究,2006,24(6):509-513.
[15] 蒲娜娜,杜国强,李明媛,等.7 种SH 系苹果砧木的AFLP分析[J].中国农学通报,2007,23(6):141-144.
[16] 胡艳青,苏 媛,韩风叶,等.嫁接黄瓜在愈合过程中的解剖观察和抗氧化酶活性的变化研究[J].内蒙古农业大学学报,2007,28(3):224-230.
[17] 黄坚钦,章滨森,陆建伟,等.山核桃嫁接愈合过程的解剖学观察[J].浙江林学院学报,2001,18(2):111-114.
[18] 孟海军.柑橘胚胎发生过程中DNA甲基化/去甲基化研究及SSR标记开发[D].武汉:华中农业大学,2006:16-25.
[19] Wassenegger M.RNA-directed DNA methylation[J].Plant Mol.Biol., 2000, 43:203-220.
[20] Xu M L, Li X Q, Korban S S.DNA-methylation alterations and exchanges during in vitro cellular differentiation in rose(Rosa hybrida L.) [J].Theor.Appl.Genet.,2004, 109(5):899-910.
[21] 陈 红,王永清.番茄与茄子嫁接接合部愈伤组织的RAPD分析[J].园艺学报,2006,33(3):565-565.
[22] 李 明,姜世玲,王幼群,等.基因转录后沉默信号可以在拟南芥嫁接体内快速双向传递[J].科学通报, 2006,51(2):142-147.
[23] Ohta Y.Graft-transformation, the mechanism for graft-induced genetic changes in higher plants [J].Euphytica,1991, 55:9l-99.
[24] Dan-Hua Z,Zhao-Huang M,Wei-Ming X,et al.Graft-induced inheritale variation in mungbean and its applocation in mungbean breeding[J].Acta Botanica Sinica,2002, 44(7):832-837.
AFLP analysis of nurse seed grafted union in Camellia oleifera during healing
FENG Jin-ling, YANG Zhi-jian, CHEN Hui
(Fujian Agriculture and Forestry University, Fuzhou 350002, Fujian, China)
To investigate whether gene changes of the nurse seed grafting union and the scion during healing of the nurse seed grafted union in Camellia oleifera.The differences of development between the nurse seed grafting unions in Camellia oleifera and the healing wound, the mother tree of the scion, the root stock and the buds from the scion were examined by AFLP(Amplified Fragment Length Polymorphism) technology.The results show that the number of bands changed relatively during the development of the nurse seed grafting union, especially at the days 4th, 16th, 29th and 35th after grafting; the change in number of bands during the development of the healing wound took place at the days 0~18th after transplantation.There were 13 specific bands in the gene among the scion, the root stock and the buds from the scion.Of them, 10 bands belonged to the buds from the scion DNA, 5 bands of which only belonged to it, the rest bands were found on both it and the root stock.The band change of the nurse seed grafting may caused by DNA methylation.The changes of the scion may resulted from DNA methylation, or affected by the pressure of the graft, may gene exchanged between the scion and the root stock.The real causes of the changes will be studied furtherly.
Camellia oleifera; nurse seed grafting; gene; AFLP
S794.4
A
1673-923X(2012)03-0141-06
2011-10-27
科技人员服务企业行动项目(2009GJC40006);福建省科技平台建设项目(2010N2001)
冯金玲(1978—),女,福建尤溪人,讲师,博士,主要从事经济林生物学研究
陈 辉(1957—),男,福建福州人,教授,博士生导师,主要从事森林培育学研究;E-mail:zjchchenh@163.com
[本文编校:谢荣秀]
