5株桃、李果实采后褐腐病菌鉴定、rDNA ITS序列与碳源代谢指纹图谱分析
2012-10-27王友升陈玉娟
王友升,张 燕,陈玉娟
(北京工商大学食品学院,食品添加剂与配料北京高校工程研究中心,北京市食品风味化学重点实验室,北京 100048)
5株桃、李果实采后褐腐病菌鉴定、rDNA ITS序列与碳源代谢指纹图谱分析
王友升,张 燕,陈玉娟
(北京工商大学食品学院,食品添加剂与配料北京高校工程研究中心,北京市食品风味化学重点实验室,北京 100048)
从采后贮藏过程中发病的桃、李果实中分离到5株丝状病原真菌。对其进行形态学特征观察以及核糖体rDNA ITS序列分析和95种碳源代谢指纹图谱分析。结果表明:5株病原真菌均为果生链核盘菌(Monilinia fructicola),其中091#和089#菌与Monilinia fructicola(FJ515894)的亲缘关系较近。5株褐腐病菌对D-果糖、麦芽糖、蔗糖等共56种碳源的代谢能力相同,包括51种最适碳源和5种不可利用碳源,其中Monilinia fructicola091#与089#的代谢指纹图谱最接近,而Monilinia fructicola086#、071#、072#的代谢指纹图谱最接近。因此利用rDNA ITS序列和碳源代谢指纹图谱均可区分桃、李果实5株褐腐病菌不同菌株之间的差异,且两种方法具有可比性。
桃果实;李果实;褐腐病菌;rDNA ITS序列分析;碳源代谢指纹图谱
桃、李属呼吸跃变型果实,采后呼吸强度大,后熟迅速,加之其皮薄多汁,采收期正值盛夏,极易受到微生物侵染,发生腐烂[1-2]。低温、气调等常见物理保鲜方法会降低果实本身的抗病力,不能有效控制销售过程中的腐烂,因而常需配合使用防腐剂。对引起果实贮藏期腐烂的主要病原微生物进行分离鉴定,是针对性筛选防腐剂,延长货架期的前提。rRNA基因内转录区间(internal transcribed spacer,ITS)分析因其可以从较短的核酸序列中获得种间及种内差异,反映生物的亲缘关系及分类关系,而广泛应用于真菌的鉴定、分类与种属间系统学研究。但ITS序列分析法用于菌种鉴定受基因库的完善程度、高度同源性序列的多少以及具体物种ITS区可变程度等因素局限[3],宜与形态特征、生长特点、生理生化指标等传统鉴定方法相结合[4]。碳源代谢指纹图谱主要反映微生物对微平板上不同碳源的利用程度,由此产的不同碳源利用模式即可表征微生物间的差异,常用于真菌的鉴定及同种突变体间的研究[5-6]。
本研究从采后贮藏期发病的桃、李果实上分离到5株病原真菌,结合形态学特征以及rDNA ITS序列分析对这5株病原菌进行鉴定,并通过构建系统进化树和碳源指纹聚类分析研究它们的亲缘关系,综合评价两种方法之间的相关性,以期为桃、李采后病原真菌的营养生理研究、病害防治及真菌种间关系研究提供参考。
1 材料与方法
1.1 菌株与培养基
菌株分离于采后贮藏过程中自然发病的‘黑琥珀’李和‘八月脆’桃果实。
PDA培养基:马铃薯200g、葡萄糖20g、琼脂18g、加水补足1L,121℃灭菌15min。ME培养基:麦芽浸粉20g、琼脂18g,加入一定量的水中,调节pH值至5.5±0.2,121℃灭菌15min。
1.2 试剂与仪器
Tris、EDTA、SDS 美国Amresco公司; DNA分子质量标准品、10×PCR Buffer、dNTP、TaqDNA聚合酶 天根生化试剂公司;无水乙醇、HCl均为分析纯。
PCR仪 美国Bio-Rad公司;高速冷冻离心机 德国Sorvall公司;生物安全柜 美国Thermo公司;微生物鉴定系统 美国Biolog公司;Axio Image A1显微镜德国Zeiss公司。
1.3 方法
1.3.1 菌种的分离、纯化与回接
分离:取发病果实病变、健康交界处组织于PDA培养基上,25℃条件下培养1~3d。
纯化:从分离菌菌落外沿取菌块,分别转接到PDA和ME培养基上,于25℃条件下培养7d,观察菌落形态和个体形态特征。
回接:挑选健康无损伤的桃、李果实,用75%的乙醇将表面消毒,然后用无菌接种针刺孔,取纯化后的菌块接种至伤口处,观察果实接种处是否出现相应病症。
1.3.2 基因组DNA的提取及ITS区PCR扩增与序列测定
用改良后SDS-氯化苄法提取基因组DNA[7]。用ITS区扩增通用引物ITS-4(5'-TCCTCCGCTTATTGATATGC-3')和ITS-5(5'-GGAAGTAAAAGTCGTAACAAGG-3')进行PCR扩增,通用引物ITS-4和ITS-5由Invitrogen公司合成。PCR产物由华大基因进行脱盐、纯化和测序,测序结果在GenBank数据库中进行同源序列搜索,并且构建系统发育进化树。
1.3.3 碳源代谢指纹图谱分析
将待测菌株在PDA培养基平板上26℃培养120h,用FF-IF浊度标准液配制菌悬液,调节T=(75±2)%。在FF微孔板中每孔加100μL菌悬液,26℃培养,每24h用Biolog微生物鉴定系统读取结果,获得病原菌的代谢指纹图谱。
2 结果与分析

图1 桃、李果实病原真菌的分离与回接Fig.1 Isolation and re-inoculation of pathogenic strains in peach and plum fruits
2.1 菌株的分离、纯化与回接由图1可知,从‘黑琥珀’李果实中分离到4株菌,其中091#和089#菌从20℃贮藏1d的果实中分离得到086#菌从1℃贮藏30d出库后并于20℃贮藏6d的果实中分离得到,071#菌分离自1℃贮藏60d出库后并于20℃贮藏6d的果实;072#菌分离自‘八月脆’桃果实。将分离得到菌株纯化后回接到健康的桃、李果实上,在接种部位均出现同样的病症,并能从该病害部位再次分离得到相应病原菌,因此,可确定它们均为桃、李果实的致病菌。
2.2 病原菌形态观察与分析
2.2.1 病原菌菌落形态观察
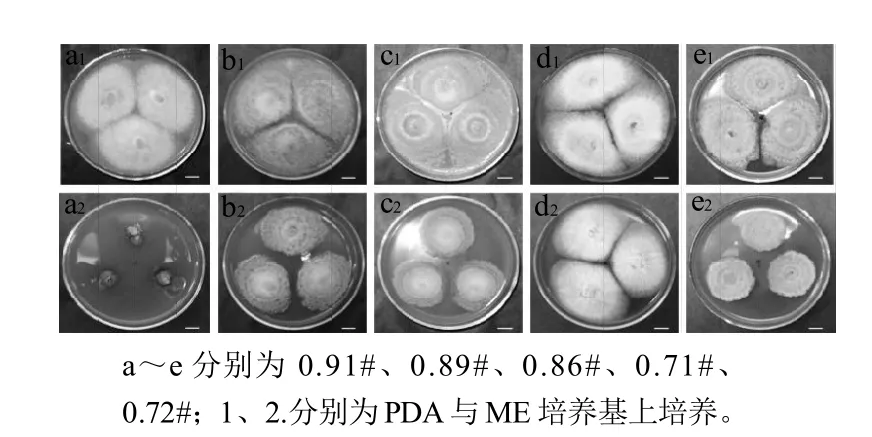
图2 桃、李果实5株病原菌在PDA和ME在25℃条件下培养7d的菌落特征Fig.2 Colony characteristics of five isolated strains cultured in ME and PDA media for 7 days at 25 ℃
由图2可知, 5株病原菌在PDA培养基上均生长旺盛,ME培养基上长势较PDA平板弱,091#菌株尤为突出,091#在PDA培养基上培养7d后,菌丝已铺满平板,浅褐色,簇生绒毛状(图2a1),而ME培养基上7d后,菌丝几乎没有扩展(图2a2)。089#、086#、072#菌落特征一致,PDA培养基上培养7d菌丝铺满平板,可以看到明显的褐色同心轮纹(图2b1、c1、e1)。ME培养基上菌丝性状同PDA板上相似,菌落直径较PDA板上小(图b2、c2、e2)。071#菌在PDA和ME培养基上均生长旺盛,7d后菌丝均铺满平板,菌丝白色致密绒毛状,菌落中央均呈现浅绿色,PDA板中央绿色更深(图 2d1、d2)。
2.2.2 病原菌显微镜观察

图3 5株病原菌的显微形态观察Fig.3 Microscopic observation of five isolated strains
由图3可知,5株病原菌显微形态一致,孢子梗具隔,顶端分枝,分生孢子单孢、串生,孢子壁光滑,孢子椭球形或柠檬形(图3a1~e1)。菌丝分枝多且内部隔膜观察明显(图a2~e2)。
2.3 基因组DNA的提取和PCR扩增
采用SDS-氯化苄法提取的5株病原菌总DNA以ITS4和ITS5为引物,经PCR扩增得到的产物经琼脂糖凝胶电泳分析和紫外检测后,可以看出所提取的DNA大小约600bp,与实验设计吻合(图4)。

图4 5株病原菌rDNA ITS电泳检测图谱Fig.4 Agarose gel electrophoresis results of rDNA ITS for five isolated strains
2.4 rDNA ITS同源性比对与系统发育分析
将分离得到的5株病原菌的rDNA ITS基因部分序列根据GenBank内登录的相应序列进行同源序列比对,通过同源性比对结果经Clustal X软件和MEGA3.1软件构建系统进化树,如图5所示。同源性比对结果表明,经BLAST比对发现,5株分离菌与Monilinia fructicola(FJ515894) ITS序列同源性皆高达99%,分别仅差1~6个碱基。将其形态特征(图2)与《真菌鉴定手册》[8]中的果生链核盘菌(Monilinia fructicola)的描述进行比较,基本一致,5株菌经鉴定均为褐腐病原菌果生链核盘菌(M. fructicola)[9]。从构建的系统进化树上可以看出,5株菌分别位于两个分支上,其中Monilinia fructicola089#、091#与Monilinia fructicola(FJ515894)在同一分枝上,Monilini afructicola071#、086#、072#在进化树的另一分支上。

图5 以rDNA ITS基因序列为分子标记的病原菌菌株系统进化树Fig.5 Phylogenetic tree of five isolated strains based on rDNA ITS sequences
2.5 碳源代谢指纹图谱分析
表1为5株病原菌对95种碳源的代谢指纹图谱,可以看出,不同菌株之间的碳源代谢指纹图谱存在一定差异,5株菌可以利用的碳源均在68种以上,共有56种利用程度相同的碳源,包括D-果糖、麦芽糖、蔗糖等51种最适碳源和5种不可利用碳源:i-赤藻糖醇、L-海藻糖、葡糖醛酰胺、γ-羟丁酸、腐胺。其中,Monilinia fructicola091#与089#的代谢指纹图谱最为相近,两者对78种碳源利用情况一致,仅对D-核糖、L-苏氨酸和α-酮戊二酸3种碳源的利用程度差异较大;与Monilinia fructicola086#代谢指纹图谱最为相近是Monilinia fructicola072#,其次是Monilinia fructicola071#。Monilinia fructicola086#与072#利用情况相同的碳源有7 8种,利用程度差异较大的碳源仅7种,Monilinia fructicola086#与071#利用情况相同的碳源有76种,利用程度差异较大的碳源仅10种。
聚类分析可将具有相似碳源代谢特征的菌进行分类,从而有效反映不同菌株在代谢上的关系。对5株Monilinia fructicola采用欧氏距离测量,每两样本间用Average Linkage法连结,以95种碳源的利用情况作为聚类变量,进行聚类分组,结果如图6所示。可以看出,通过系统聚类分析可将5株病原真菌直观清楚的区分开来,其中Monilinia fructicola091#与089#、Monilinia fructicola086#与072#首先聚为一类,然后Monilini afructicola071#与086#、072#相聚。

表1 5株褐腐病菌的碳源代谢指纹图谱Table 1 Carbon source metabolic fingerprinting of five isolated strains

图6 5株褐腐病菌的碳源特征聚类分析Fig.6 Clustering analysis of five isolated strains by metabolic fingerprints
3 讨 论
已报道[10]桃、李果实贮藏期的侵染性病害包括褐腐病、根霉病、青霉病、孢霉病和灰霉病等。本研究从采后贮藏过程的发病桃、李果实上多次分离纯化病原菌,虽然也可分离出串珠状赤霉(G i b b e r e l l a moniliformis)、灰葡萄孢(Botrytis elliptica)、草酸青霉(Penicillium oxalicum)等病原菌(结果待发表),但分离到最多是果生链核盘菌(Monilinia fructicola)。由果生链核盘菌引起的褐腐病是桃、李、樱桃等核果类果实采后和贮藏期的主要病害[11],该菌可以侵染未成熟的果实,在贮藏期发病,菌体生长迅速,极易扩散[12],常引起贮藏、运输和销售中的大量烂果,损失严重。
侵染性病害的发病过程常受到寄主自身营养条件的限制。蔗糖、山梨醇、果糖、葡萄糖、苹果酸和奎宁酸是桃、李果实中的主要糖酸组分[13-14],同时也是分离到的5株果生链核盘菌的最适碳源(表1)。所以桃、李果实自身的营养特性可能是导致褐腐病成为其贮藏期主要病害的原因。同时,碳源代谢图谱表明,5株菌可以利用的碳源均在68种以上,可见果生链核盘菌对碳源的适应性非常强,这可能是褐腐病侵染范围广,难以防治的主要原因。
ITS序列进化速率相对较快,可以提供较丰富的变异位点和信息位点,近年来被广泛用于解决不同地理分布、种间、种内甚至居群间被子植物、低等动物系统发育和分类问题[15-16]。本研究分离到的5株果生链核盘菌的ITS区序列具有一定的变异性,这种变异可能属于种内变异(图5)。由系统进化树的结果可以看出,Monilinia fructicola091#跟089#的亲缘关系较近,而Monilinia fructicola086#、071#、072#的亲缘关系较近。碳源代谢指纹分析也可以看出,M o n i l i n i a fructicola091#与089#代谢模式相近,Monilinia fructicola086#、071#、072#菌代谢模式相近,这与系统进化树分析所揭示的5株果生链核盘菌间的亲缘关系吻合,表明果生链核盘菌之间的遗传差异同样可体现在它们的碳源代谢特征上。本课题组前期对3株罗伦隐球酵母的26S rDNAD1/D2区序列和代谢指纹图谱比较,也发现碳源代谢可以在一定程度上反映亲缘关系[17]。然而,与进化树结果相比,碳源代谢指纹图谱分析还可以将Monilinia fructicola086#、071#、072#的亲缘关系进一步细分,即Monilinia fructicola086#与072#关系最近,其次是Monilinia fructicola071#。因此,碳源代谢指纹图谱分析可以为种间变异、种内系统学研究及种下等级划分提供新的佐证。
4 结 论
从采后贮藏期自然发病的桃、李果实上分离得到5株丝状真菌,经分子生物学结合形态学鉴定均为果生链核盘菌(Monilinia fructicola)。通过rDNA ITS序列分析可以看出,5株果生链核盘菌的ITS序列具有一定的变异性。对95种碳源代谢指纹图谱的分析表明,5株菌可以利用的碳源均在6 8种以上,其中M o n i l i n i a fructicola091#与089#的碳源代谢指纹相近,而Monilinia fructicola086#、071#与072#的碳源代指纹相近。利用rDNA ITS序列和碳源代谢指纹图谱均可区分桃李果实5株褐腐病菌不同菌株之间的差异,且两种方法具有可比性。
[1] 郭晓敏, 安琳, 王友升, 等. 两种贮藏温度下 “ 黑琥珀”李果实品质及不同部位抗氧化活性的变化规律[J]. 食品科学, 2010, 31(20): 425-429.
[2] GIRARDI C L, CORRENT A R, LUCCHETTA L, et al. Effect of ethylene, intermittent warming and controlled atmosphere on postharvest quality and the occurrence of woolliness in peach (Prunus persicacv.Chiripá) during cold storage[J]. Postharvest Biology and Technology,2005, 38(1): 25-33.
[3] 燕勇, 李卫平, 高雯洁, 等. rDNA-ITS序列分析在真菌鉴定中的应用[J]. 中国卫生检验杂志, 2008, 18(10): 1958-1961.
[4] 吴群, 姬可平. rRNA基因内转录间隔区作为遗传标记的应用现状及前景[J]. 时珍国医国药, 2008, 19(11): 2806-2808.
[5] 王强, 戴九兰, 吴大千, 等. 微生物生态研究中基于BIOLOG方法的数据分析[J]. 生态学报, 2010, 30(3): 817-823.
[6] SINGH M P. Application of Biolog FF MicroPlate for substrate utilization and metabolite profiling of closely related fungi[J]. Journal of Microbiological Methods, 2009, 77(1): 102-108.
[7] PRYOR B M, GILBERTSON R L. Molecular phylogenetic relationships amongstAlternariaspecies and related fungi based upon analysis of nuclear ITS and mt SSU rDNA sequences[J]. Mycological Research,2000, 104(11): 1312-1321.
[8] 魏景超. 真菌鉴定手册[M]. 上海: 上海科学技术出版社, 1979: 566-614.
[9] LANDEWEERT R, LEFFLANG P, KUYPER T W, et al. Molecular identification of ectomycorrhizal mycelium in soil horizons[J]. Applied and Environmental Microbiology, 2003, 69(1): 327-333.
[10] 郑重禄. 李果的贮藏特性与保鲜技术[J]. 中国果蔬, 2000(1): 21.
[11] BOEHM E W, MA Z, MICHAILIDES T J. Species-specific detection ofMonilinia fructicolafrom california stone fruits and flowers[J].Phytopathology, 2001, 91(5): 428-439.
[12] GONC F P, MARTINS M C, GERALDO J, et al. Postharvest control of brown rot andRhizopusrot in plums and nectarines usingcarnaubawax[J]. Postharvest Biology and Technology, 2010, 58: 211-217.
[13] 胡玲, 王友升, 李丽萍. GC-MC法同时测定 “ 安哥诺”李果实中非挥发性物质衍生化条件研究[J]. 食品科学, 2010, 31(14): 186-190.
[14] 李艳萍, 牛建新, 陈清. 桃果实中糖酸物质代谢的影响因素研究进展[J]. 植物生理科学, 2007, 23(8): 212-216.
[15] 林珊, 郑伟文, 吴锦忠, 等. 不同来源莲rDNA ITS的PCR扩增、克隆及序列分析[J]. 中国中药杂志, 2007, 32(8): 671-675.
[16] 王晓玲, 郭安平, 彭于发, 等. 海南普通野生稻不同居群rDNA ITS区序列的比较分析[J]. 热带作物学报, 2008, 29(4): 478-484.
[17] 姚子鹏, 郭晓敏, 王友升, 等. 3株罗伦隐球酵母的26S rDNAD1/D2区序列、代谢指纹图谱及生防效力比较分析[J]. 食品科学, 2011,32(1): 135-140.
Identification, rDNA ITS Analysis and Carbon Metabolic Fingerprinting of FiveMonilinia fructicolaStrains Isolated from Postharvest Peach and Plum Fruits
WANG You-sheng,ZHANG Yan,CHEN Yu-juan
(Beijing Higher Institution Engineering Research Center of Food Additives and Ingredients, Beijing Key Laboratory of Food Flavor Chemistry, School of Food and Chemical Engineering, Beijing Technology and Business University, Beijing 100048, China)
In the present study, four pathogenic strains named as 091, 089, 086 and 071, respectively were isolated from infected black amber plum, and one pathogenic strain named as 072 from infected Bayuecui peach during postharvest storage. Morphological observation and rDNA ITS analysis indicated that all five pathogenic strains wereMonilinia fructicola. The result obtained also revealed that strains 091 and 089 had a close phylogenetic relationship with Monilinia fructicola (FJ515894). Metabolic fingerprinting suggested that these five strains had the same ability to metabolize 56 carbon sources including 51 optimal ones and 5 unutilized ones.Monilinia fructicola091 and 089 shared the most similar metabolic fingerprints, whereasMonilinia fructicola086, 071 and 072 were the most similar. These studies demonstrated that both rDNA ITS analysis and carbon metabolic fingerprinting allows distinguishing among theMonilinia fructicolastrains isolated and may be considered as being comparable.
peach fruit;plum fruit;Monilinia fructicola;rDNA ITS sequence analysis;carbon metabolic fingerprinting
Q93-331
A
1002-6630(2012)16-0246-05
2012-04-11
国家“863”计划项目(2012AA101607);国家自然科学基金青年科学基金项目(30901009);北京市自然科学基金项目(6122003)
王友升(1976—),男,副教授,博士,研究方向为食品生物技术。E-mail:wangys@th.btbu.edu.cn
