钙对低温下山定子根系活力及氮代谢的影响
2012-10-19张宝贵梁启伍韩艺智史婧贤
梁 立,张宝贵 ,梁启伍,韩艺智 ,史婧贤
(1.天津市宝坻区苗圃场,天津 宝坻 301800;2.天津市宝坻区林业技术推广站,天津 宝坻 301800;3.天津市宝坻区森保站,天津 宝坻 301800)
氮代谢是高等植物最重要的代谢过程之一,它直接影响树体对氮素的吸收和同化,进而影响果实产量和品质。近年来,关于低温条件下添加钙素对细胞保护酶活性以及缓解低温胁迫的研究很多,但有关钙对低温条件下氮代谢功能影响的研究较少。因此,本试验以北方地区常用苹果砧木山定子为试材,研究低温条件下钙对根系氮代谢的影响。
1 材料与方法
1.1 试验材料与试验设计
试验于2009年3~7月进行,山定子种子经层积催芽后,播种到育苗基质中,待长到5片真叶时挑选生长一致的幼苗移入装有基质的营养钵(12 cm×13 cm)中,待根系充满营养钵时,小心冲洗出根系进行水培试验。水培试验于人工气候箱中进行,培养温度为 25℃(昼)和 15℃(夜),光照强度 400 μmol·m-2·s-1,光照时间 12 h,相对湿度70%~80%。处理营养液以Hoagland全营养液为基础(其中钙浓度为 4 mmol·L-1),设计 4个钙浓度处理 Ca1、Ca2、Ca3、Ca4,分别添加 4 mmol·L-1、8 mmol·L-1、12 mmol·L-1、16 mmol·L-1硝 酸 钙[Ca(NO3)2]。各处理分别水培一周后转入低温处理,处理温度为10℃(昼)和2℃(夜),以始终处在常温25℃(昼)和15℃(夜)的植株为对照,分别于低温处理3 d后取样测定各项指标,每处理10株,所有试验均重复3次。
1.2 根系活力及氮代谢相关物质测定方法
根系活力采用TTC染色法测定,蛋白质含量采用考马斯亮蓝法测定,游离氨基酸含量采用茚三酮比色法测定,游离氨基酸种类采用5%磺基水杨酸提取,滤膜过滤后用日立L8800型氨基酸自动分析仪测定。
1.3 根系氮代谢关键酶活性测定方法
硝酸还原酶采用磺胺比色法测定。取根系0.5 g,加入10 ml磷酸缓冲液,真空充气10 min,中间放气2~4次,置于暗室20 min,取出后加入30%三氯乙酸1 ml,立即振荡,取浸提液2 ml置于试管中,加入1%的磺胺试剂4 ml和0.02%的α-萘胺试剂4 ml,振荡摇匀静置 30 min,在540 nm下测定其吸光度。
按Zhang CF等介绍的方法在根系中提取粗酶液,每克鲜样加入3 ml提取缓冲液。粗酶液用于 GS、NADH-GOGAT、NADH-GDH 活性测定。
GS活性参照Rhodes等的方法测定,一个GS活性单位定义为该反应条件下,每分钟催化形成1μmol的 γ-谷氨酰异羟肟酸 (γ-glutamy-lhydroxamate)所需要的酶量。NADH-GOGAT活性采用Singh和Srivastava的方法测定,以每分钟反应混合液于30℃减少1μmol的NADH为一个酶活性单位。总NADH-GOGAT活性计算以每分钟每毫克酶液催化NADH减少多少μmol表示。NADH-GDH活性参照Loulakakis和Roubelakis的方法测定,GDH活性以每分钟在30℃下氧化1μmol的NADH为一个酶活性单位。
1.4 呼吸速率的测定
参照Bouma和毛志泉等的方法进行测定,仪器采用英国HANSATECH公司生产的液相Oxy-Lab型氧电极。根系测定时取直径1.5 mm左右,长度约2~3 cm的吸收根,用双面刀将根切成2 mm左右根段(0.05 g),迅速称取0.05 g放入反应杯,加盖并启动测量程序。反应杯中反应液温度用恒温浴控制在25℃,每个处理3个(株)重复。
1.5 呼吸途径的测定
参照余让才和潘瑞炽的方法进行测定。糖酵解途径(EMP)、三羧酸循环(TCA)和磷酸戊糖途径(PPP)分别用 0.5 mol·L-1的 NaF、丙二酸和 Na3PO4抑制,反应介质为0.2 mol·L-1pH6.8的磷酸缓冲液。每个途径测定3次。
2 结果与分析
2.1 钙对低温下山定子根系活力的影响
如图1所示,低温使山定子根系活力降低,与常温对照差异显著,添加外源钙后山定子根系活力有所增加,添加4 mmol·L-1钙时根系活力高于对照,与对照没有显著差异,添加8 mmol·L-1钙时根系活力虽低于常温对照,但仍高于低温对照,当外源钙浓度增加到 12 mmol·L-1和 16 mmol·L-1时根系活力降低,与对照呈显著差异。
2.2 钙对低温下山定子根系氮代谢关键酶活性的影响
如图2所示,低温降低了山定子根系NR活性,但与常温相比没有明显差异。添加外源钙能提高根系NR活性,四个钙浓度处理根系NR活性均高于低温对照和常温对照,添加4 mmol·L-1钙处理与低温对照呈显著差异,但各个浓度钙处理与常温对照均无显著差异。
低温明显增加了山定子根系GS活性,与相同处理的常温对照呈显著差异。添加外源钙与常温对照相比均提高了根系GS活性,且增加的幅度与钙浓度成正比。但添加外源钙处理与同温度的对照相比在低浓度时降低了根系GS活性,而高浓度时(16 mmol·L-1)GS 活性略有增加。 添加4 mmol·L-1钙和 8 mmol·L-1钙处理与同温度的对照呈显著差异;除添加4 mmol·L-1钙处理外均与常温对照差异显著。
由图2可知,低温显著降低了山定子根系NADH-GOGAT活性,与相同处理的常温对照差异显著。添加不同浓度外源钙对根系NADHGOGAT活性的影响不同,但根系NADH-GOGAT活性均低于常温对照;与低温对照相比,添加4 mmol·L-1钙、 添加 12 mmol·L-1钙和 16 mmol·L-1钙处理根系NADH-GOGAT活性有所增加,但添加 8 mmol·L-1钙处理根系 NADH-GOGAT活性降低,添加外源钙处理与相同温度的对照相比均没有显著差异,但添加8 mmol·L-1钙和添加12 mmol·L-1钙处理根系NADH-GOGAT活性与常温对照差异显著。

图1 钙对低温下山定子根系活力的影响
低温降低了山定子根系NADH-GDH活性,与相同处理的常温对照没有明显差异。外源钙影响根系NADH-GDH活性,添加4 mmol·L-1钙和添加8 mmol·L-1钙时根系NADH-GDH活性高于低温对照,但没有显著差异,当钙浓度增加时根系NADH-GDH活力明显降低,与其他处理差异显著。
2.3 钙对低温下山定子根系GS/GDH比值的影响
如图3所示,低温和外源钙均未改变山定子根系氮代谢途径。低温后根系GS/GDH比值增加,与常温对照差异显著。外源钙在低浓度内降低了根系GS/GDH比值,与低温对照呈显著差异,而与常温对照没有明显差异;但钙浓度增加后GS/GDH比值迅速增加,与低温对照差异显著。
2.4 钙对低温下山定子根系氮代谢相关物质的影响
由图4可知,低温降低了山定子根系可溶性蛋白质含量,与常温对照呈显著差异。添加外源钙处理与相同温度的对照相比增加了山定子根系可溶性蛋白质含量,其中4 mmol·L-1钙处理与低温对照呈显著差异,其余处理与低温对照没有明显差异。低温加钙处理可溶性蛋白质含量均低于常温对照。
由图5可知,低温降低了山定子根系游离脯氨酸含量,但与常温对照没有显著差异。添加钙处理增加了根系游离脯氨酸含量,其中添加4 mmol·L-1钙处理和 8 mmol·L-1钙处理与低温对照和常温对照均差异显著。
2.5 钙对低温下山定子根系呼吸速率的影响
如图6所示,低温降低了山定子根系总呼吸速率,与常温对照相比呈显著差异。外源加钙处理改变了根系总呼吸速率,4 mmol·L-1钙处理和8 mmol·L-1钙处理增加了根系总呼吸速率,与低温对照呈显著差异;而高浓度钙处理使根系总呼吸速率降低,与低温对照没有显著差异,但与常温对照差异显著。
2.6 钙对低温下山定子根系呼吸途径的影响
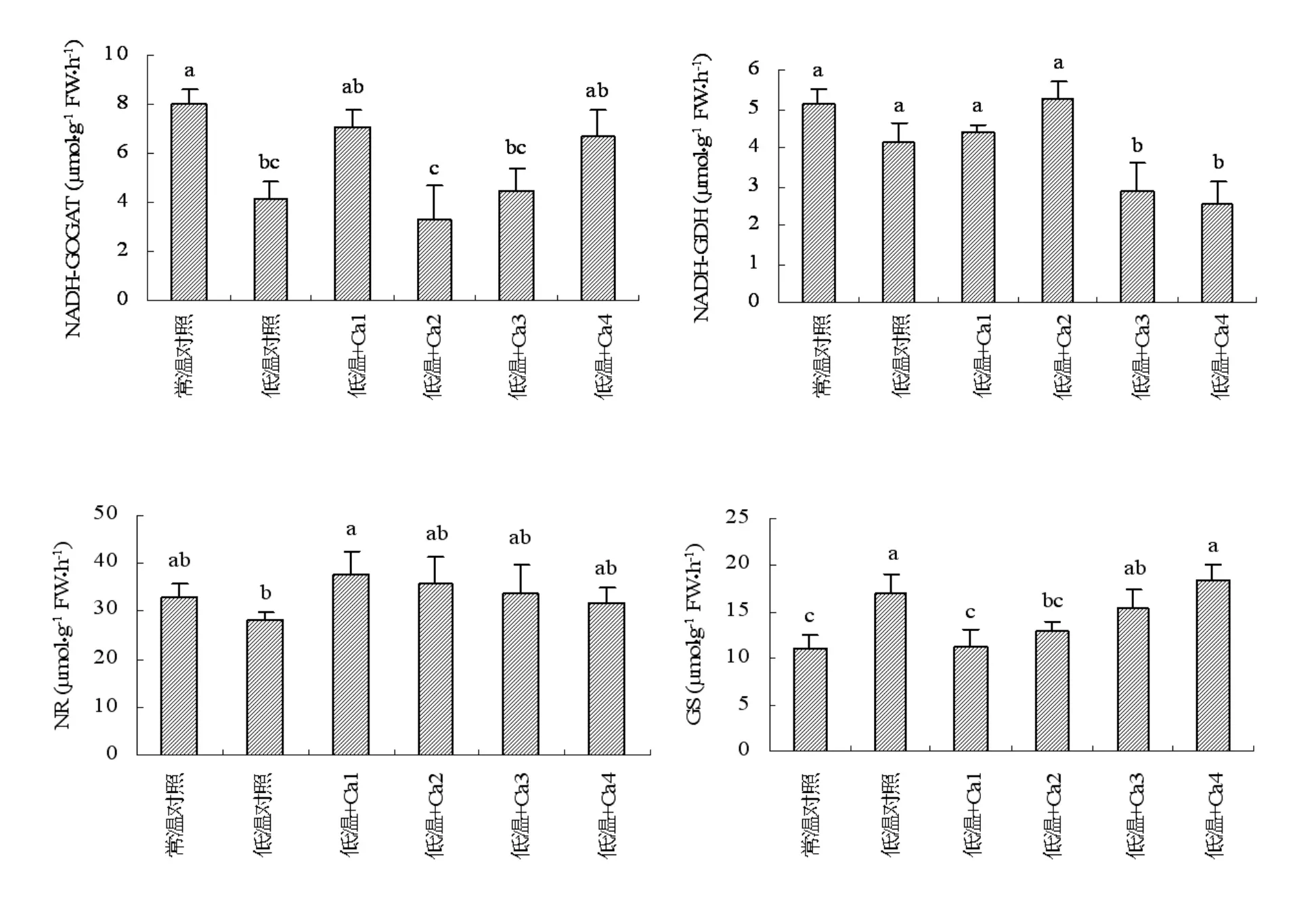
图2 小分子有机物对低温下山定子根系氮代谢关键酶活性的影响
由图7可知,低温和外源钙均未改变山定子根系呼吸途径,山定子根系呼吸仍然以由EMP进入TCA途径为主。低温处理后TCA途径所占比例明显降低,而EMP途径所占比例增加,均与对照呈显著差异。添加外源钙素后,EMP途径所占比例增加,但TCA途径所占比例有所降低,使EMP途径明显高于TCA途径和PPP途径所占比例,从而使EMP进入TCA和PPP受阻。添加4 mmol·L-1钙处理、8 mmol·L-1钙处理和 12 mmol·L-1钙处理根系PPP途径所占比例均低于低温对照,且 4 mmol·L-1钙处理和 8 mmol·L-1处理与低温对照呈显著差异。
3 结论与讨论
3.1 钙对低温条件下根系氮代谢的影响
钙是植物体必需的营养元素之一,它对果树氮代谢、生长发育、果实品质、抗病性的影响已有广泛报道。Juan etal.的研究发现番茄根系NR、NiR、GS、GOGAT、PEPC 活性随溶液中钙浓度的提高而增加。鲁翠涛等研究也表明,不同浓度Ca2+培养后,小麦叶片含Ca2+/CaM-PK的激酶活性上升趋势与NR、GS活性上升趋势一致,但当Ca2+浓度达到1 mmol·L-1后,酶活性不再持续升高,说明依赖于Ca2+/CaM-PK的蛋白激酶对NR和GS具有一定的调节作用。近年来研究表明,钙对低温的响应很敏感,低温刺激后细胞中有个Ca2+浓度迅速升高的过程,此过程在时间、机制上和冷胁迫感应过程紧密联系,因此,人们推测Ca2+作为第二信使参与了植物低温信号传导,同时Ca2+还参与植物冷驯化应答和快速冷耐受反应,提高植物对低温的抗逆性。本试验结果表明,添加外源钙对山定子根系氮代谢的影响因钙浓度不同而有差异,添加低浓度钙可以提高低温下根系活力、NR、NADH-GOGAT和 NADH-GDH活性、可溶性蛋白质含量和游离脯氨酸含量,同时降低根系GS活性和GS/GDH比值,而添加高浓度钙的结果与之相反。低浓度钙对根系氮代谢活性的影响基本抵消了低温引起的氮代谢关键酶活性变化,使之接近常温对照的水平,对低温下根系氮代谢功能有明显的改善作用,而高浓度钙处理则表现出对氮代谢的抑制作用,这与闫童等的研究结果一致,可能是钙离子作为渗透调节物质对根系起保护作用,而高浓度钙可能导致膜质过氧化而引起氮代谢紊乱。
3.2 钙对低温条件下根系呼吸作用的影响
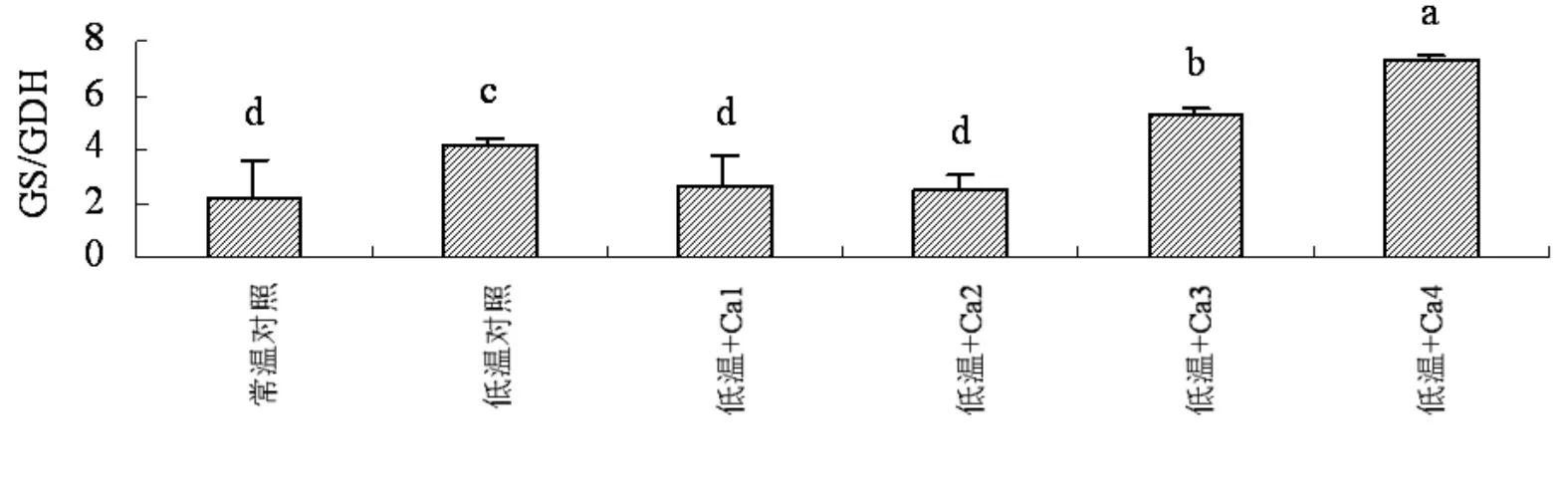
图3 钙对低温下山定子根系GS/GDH比值的影响

图4 钙对低温下山定子根系可溶性蛋白的影响

图5 钙对低温下山定子根系游离脯氨酸的影响

图6 钙对低温下山定子根系总呼吸速率的影响
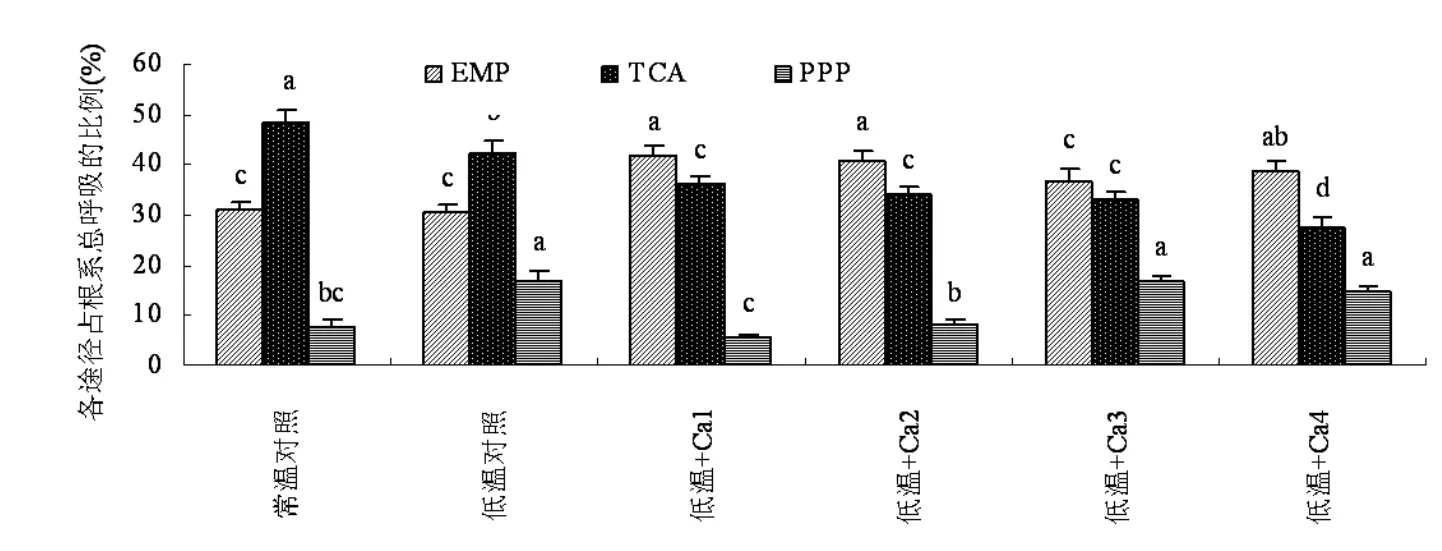
图7 钙对低温下山定子根系呼吸途径的影响
钙对低温下根系呼吸速率的影响也表现出低促高抑的作用效果。同时外源钙处理改变了山定子根系呼吸途径,从以TCA途径为主转为以EMP途径为主,且随着钙浓度的提高TCA途径所占比例不断降低而PPP途径所占比例增加。研究表明外源钙可以提高低氧和淹水胁迫下根系呼吸代谢关键酶活性,但关于低温条件下钙对呼吸速率和呼吸途径影响的研究还未见报道,具体机制有待于进一步研究。
[1]Zhang C F,Peng S B,Peng X X,et al.Response of glutamine synthetase isoforms to nitrogen sources in rice(Oryza sativa L.)roots.Plant Science[J].1997,(125)∶163-170.
[2]毛志泉,王丽琴,沈向,等.有机物料对平邑甜茶实生苗根系呼吸强度的影响 [J].植物营养与肥料学报,2004,10(2)∶171-175.
[3]余让才,潘瑞炽.蓝光对水稻幼苗呼吸代谢的影响[J].中国水稻科学,1996,10(3)∶159-162.
