色氨酸对5~10周龄扬州鹅生长性能和屠宰性能的影响
2012-10-19太仓市畜牧兽医站魏宗友
太仓市畜牧兽医站 魏宗友
扬州大学动物科学与技术学院 王洪荣*
色氨酸(Trp)是动物体内含量最低的必需氨基酸,同时也是家禽第三或第四限制氨基酸。在Trp缺乏的饲粮中,添加适量Trp能够提高畜禽的生长性能和屠宰性能。近年来,国内外就Trp对肉鸡、蛋鸡(饶巍等,2011)和鸭(王勇生等,2005)生长性能和屠宰性能方面进行了大量研究(周斌等,2011;张括等,2011),而Trp对鹅的研究鲜见报道。因此,本试验以饲喂方式和Trp水平为因子,利用2×3完全随机因子设计,研究饲粮中不同Trp水平对于扬州鹅生长性能和屠宰性能的影响。
1 材料与方法
1.1 试验动物与饲养管理 选取同一批出雏、体质健壮、体重接近、饲养管理一致的21日龄扬州公鹅270只,随机分成6组,每组3个重复,每个重复15只鹅。试验鹅经过7 d逐渐过渡为正式试验饲粮,29日龄开始正式试验,为期49 d。供试雏鹅采用舍内地面平养,自由饮水,控制舍内温度、湿度、光照,定期清洁鹅舍及周围环境,其他饲养管理以及有关免疫程序按照常规方法进行。
1.2 试验设计 试验采用2×3二因子完全随机设计,饲喂方式分别为自由采食和限制饲喂,限制饲喂组鹅饲料供给量为前一天自由采食组饲料消耗量的90%。色氨酸设3个水平,在基础饲粮中分别以低(0%)、中(0.08%)、高(0.16%)3 个水平添加L-Trp(纯度为99%,德固赛公司提供),各组饲粮中Trp含量分别为0.14%、0.22%和0.30%。基础饲粮组成及营养水平见表1。
1.3 测定指标及方法
1.3.1 生长性能 在正式试验的第6周 (70日龄)早上7∶00对扬州鹅逐只空腹称重 (停饲12 h),以重复为单位记录试验鹅的平均日采食量(ADFI),并计算平均日增重(ADG)和饲料转化率(FCR)。
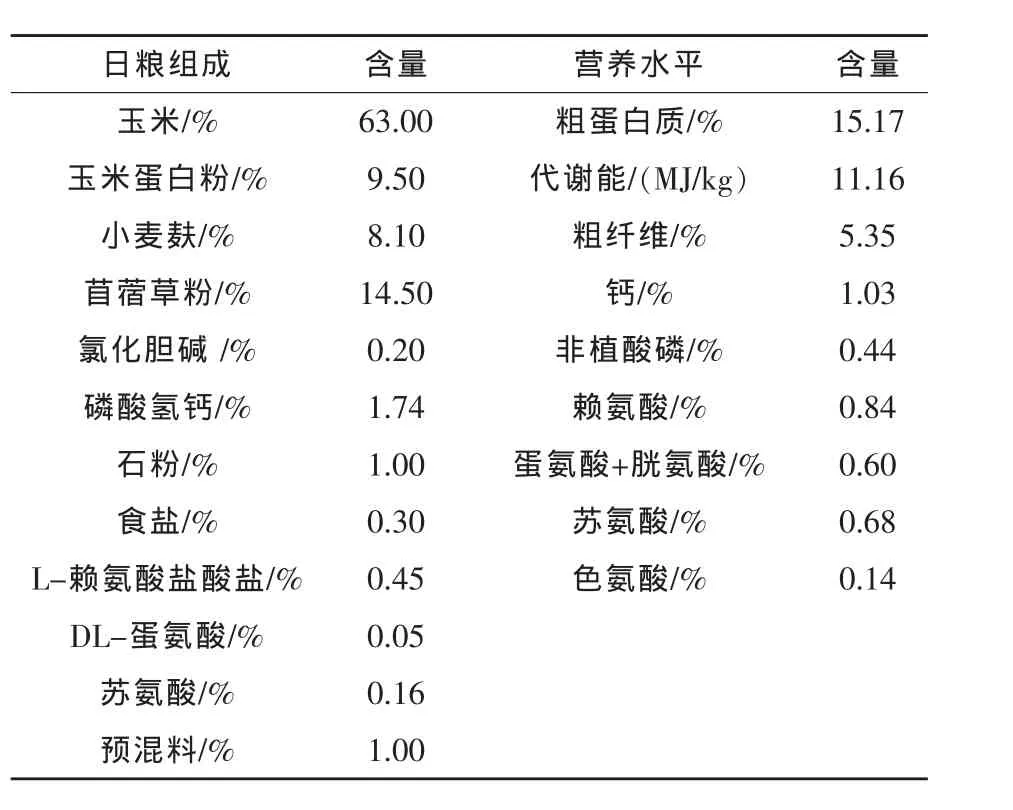
表1 基础日粮组成及营养水平(风干基础)
1.3.2 屠宰性能 在正式试验的第6周 (70日龄)清晨8∶00,每个重复组选取3只平均体重接近的鹅屠宰。宰前停饲12 h,然后称取活重、放血、拔毛,分别测定屠体重、全净膛重、半净膛重、腹脂重、胸肌重和腿肌重,并计算屠宰率、全净膛率、半净膛率、腹脂率、胸肌率和腿肌率。试验操作参照杨宁(2002)方法进行。
1.4 统计分析 试验数据采用SAS 9.1.3软件中的ANOVA模块进行两因素方差分析,运用Duncan’s法进行多重比较,P<0.05为差异显著。统计分析模型为:
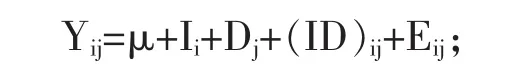
式中,Yij为性状测定值,μ为群体均值,Ii为饲喂方式效应,Dj为色氨酸效应,(ID)ij为饲喂方式和色氨酸因素的交互效应,Eij为随机误差效应。
2 结果与分析
2.1 饲喂方式和色氨酸水平对扬州鹅生长性能的影响 由表2可知,28~42日龄自由采食组扬州鹅ADFI和ADG极显著高于限制饲喂组 (P<0.01),FCR 极显著低于限制饲喂组(P<0.01)。与低Trp组相比,中Trp组扬州鹅ADFI显著提高8.04%(P<0.05),高色氨酸组提高3.50%(P>0.05)。中Trp组ADG较低Trp组和高Trp组显著提高 10.21%和 8.15%(P<0.05),而 Trp对 FCR无显著影响(P>0.05)。就交互作用而言,饲喂方式和Trp水平对28~42日龄扬州鹅ADFI、ADG和FCR有交互作用(P<0.05)。

表2 饲喂方式和色氨酸水平对28~42日龄扬州鹅生长性能的影响
由表3可知,42~56日龄扬州鹅自由采食组ADFI和FCR极显著高于限制饲喂组(P<0.01),ADG显著低于限制饲喂组 (P<0.05)。与低Trp组相比,中Trp组42~56日龄扬州鹅ADFI显著提高 5.92%(P<0.05), 高 Trp组 ADFI提高2.44%(P>0.05)。Trp水平对鹅ADG和FCR无显著影响(P>0.05)。就交互作用而言,饲喂方式和Trp水平对42~56日龄扬州鹅ADFI有交互作用(P<0.05),而对ADG和FCR不存在交互作用(P>0.05)。

表3 饲喂方式和色氨酸水平对42~56日龄扬州鹅生长性能的影响
由表4可知,56~70日龄自由采食组扬州鹅ADFI极显著高于限制饲喂组(P<0.01),ADG显著高于限制饲喂组(P<0.05),饲喂方式对FCR无显著影响(P>0.05)。Trp水平对56~70日龄扬州鹅 ADFI、ADG和 FCR无显著影响 (P>0.05)。就交互作用而言,饲喂方式和Trp水平对56~70日龄扬州鹅ADG和FCR有交互作用(P<0.05),而对 ADFI不存在交互作用(P>0.05)。
由表5可知,28~70日龄自由采食组扬州鹅ADFI和ADG极显著高于限制饲喂组(P<0.01),而饲喂方式对FCR无显著影响(P>0.05)。与低Trp组和高Trp组相比,中Trp组扬州鹅ADFI显著提高5.17%和3.91%(P<0.05)。中Trp组扬州鹅ADG较低Trp组和高Trp组显著提高8.33%和8.36%(P<0.05),但Trp水平对FCR无显著影响(P>0.05)。就交互作用而言,饲喂方式和Trp水平对28~70日龄扬州鹅ADFI和ADG有交互作用(P<0.05),而对FCR不存在交互作用(P>0.05)。
2.2 饲喂方式和色氨酸水平对扬州鹅屠宰性能的影响 由表6可知,自由采食组扬州鹅全净膛率、胸肌率和腹脂率显著高于限制饲喂组 (P<0.01),而饲喂方式对屠宰率、半净膛率和腿肌率则无显著影响(P>0.05)。中Trp组和高Trp组扬州鹅胸肌率较低Trp组分别提高15.23%和11.04%,而Trp水平对扬州鹅屠宰率、全净膛率、半净膛率、腿肌率和腹脂率无显著影响 (P>0.05)。饲喂方式和Trp水平对70日龄扬州鹅腹脂率有交互作用(P<0.01),而对其他各项屠宰性能指标无显著影响(P>0.05)。
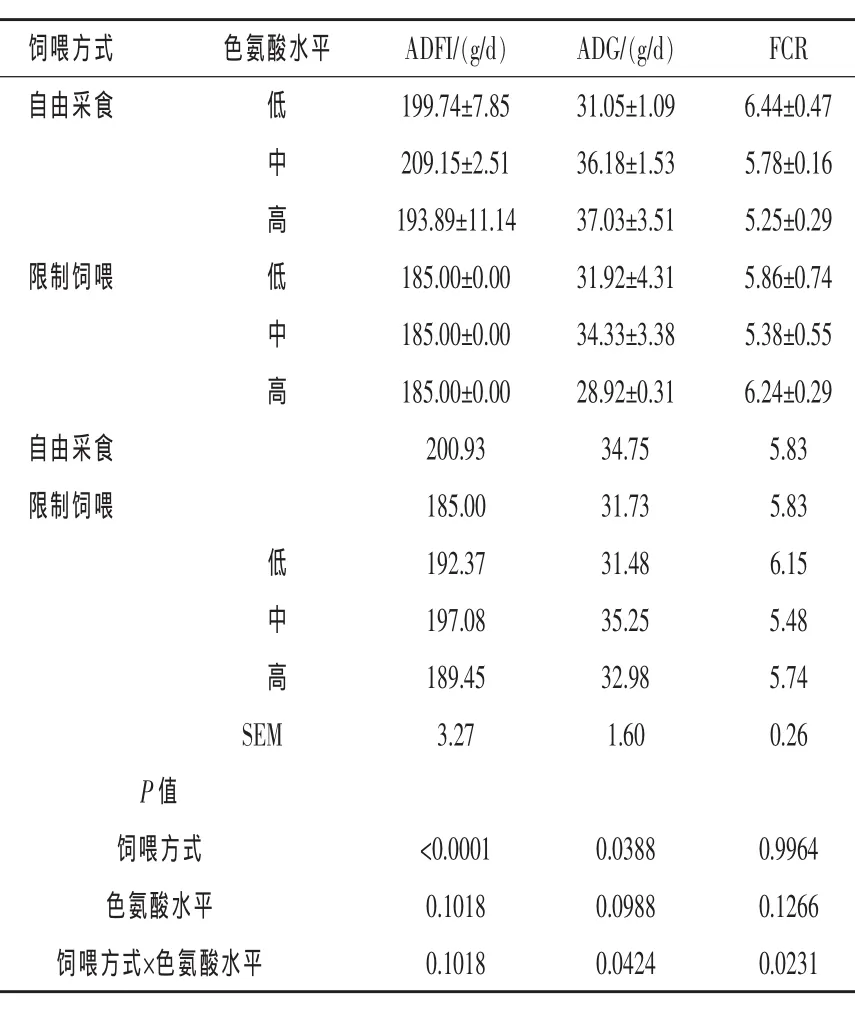
表4 饲喂方式和色氨酸水平对56~70日龄扬州鹅生长性能的影响

表5 饲喂方式和色氨酸水平对28~70日龄扬州鹅生长性能的影响
3 讨论

表6 饲喂方式和色氨酸水平对70日龄扬州鹅屠宰性能的影响%
3.1 色氨酸对扬州鹅采食量的影响 Trp是维持动物生长的必需氨基酸,对动物的采食量有重要的影响。相对于Trp含量较低的饲粮,动物会选择性地采食高Trp水平的饲粮。Harms和Russell(2000)在蛋鸡上研究发现,随着Trp水平从0.12%增加到0.20%,商品代蛋鸡的采食量也呈现显著增加的趋势。Shan等(2003)研究报道,肉鸡分别饲养在室温25℃和高温35℃条件下时,在Trp含量为0.09%的玉米-玉米蛋白粉型饲粮中添加不同水平的Trp,肉鸡采食量均显著增加。Corzo等 (2005b)、Andrew 等 (2005) 和 Rosa等(2001)均同样研究发现,缺乏Trp会抑制肉仔鸡采食量,在饲粮中添加Trp以后,采食量呈现不同程度的增加。本试验结果表明,当采食低Trp水平饲粮时,扬州鹅采食量显著降低,适当添加Trp可以改善这一状况,使得采食量增加,但Trp含量过高时,结果采食量反而呈现下降的趋势,这与以往的研究结果基本一致。结果说明,饲粮中适当添加色氨酸可提高动物采食量,Trp含量过高、过低均会对动物采食量产生一定程度的抑制作用。
色氨酸作为5-HT的前体物,经过羟化酶、脱羧酶的作用生成5-HT,其参与畜禽采食量的调节可能是由5-HT调控来发挥作用的。5-HT是一种经典的神经递质,具有广泛的生物学功能,参与机体摄食、免疫等生理功能的调节。席鹏彬等(2009)研究表明,与0.11%Trp缺乏组相比,0.14%~0.23%Trp添加组公、母鸡的日采食量分别提高4.5%~19.5%和2.2%~9.0%,下丘脑5-HT浓度分别提高37.9%~83.6%和7.3%~38.7%;利用相关分析得出,63日龄黄羽肉鸡血清游离Trp浓度(R2=0.94,P=0.018)或下丘脑 5-HT 浓度(R2=0.95,P=0.015)与其采食量呈显著正相关;这说明饲粮中Trp可能是通过下丘脑5-HT神经递质的作用来调节黄羽肉鸡采食量。然而Rosebrough(1996)研究发现,饲粮中Trp含量过高,会抑制肉鸡的采食量,大脑中5-HT水平则随着饲粮Trp水平的增加而增加。Denbow等(1993)在火鸡上发现,Trp可显著提高5-HT含量,但对采食量无影响。席鹏彬等(2009)的研究结果与上述不同,原因可能是由于环境、物种、Trp添加水平、年龄等因素所致,其具体的作用机理还有待于进一步探讨。
3.2 色氨酸对扬州鹅平均日增重的影响 在常规饲料原料中,Trp分别是玉米和大豆粕的第二和第三限制性氨基酸。近年来,由于在配合饲料中大量使用合成的Lys和Met,使得Trp成为饲料中主要的限制性氨基酸。大量研究表明,饲粮中适当添加Trp可明显促进动物生长 (饶巍等,2011;周斌 等 ,2011;Emadi 等 ,2010;Corzo 等 ,2005a、b;Fatufe等,2005)。然而这种促生长的作用是由于Trp本身的作用还是由于采食量的增加而导致,到目前为止有关饲喂方式和Trp水平对家禽的影响仍鲜见报道。本研究表明,与限制饲喂相比,自由采食显著提高了扬州鹅的平均日增重,高Trp组和低Trp组的平均日增重差异不显著,而中Trp组扬州鹅的平均日增重显著高于低Trp组,Trp水平对于饲料转化率无显著影响。而Page等(1993)、Adeola 和 Ball(1992)研究表明,在猪饲料中过量添加晶体Trp对其生长性能不会产生有益的影响。结合本研究可以发现,动物采食适当水平的Trp可以提高生长性能,这可能是由于氨基酸的平衡所致。动物采食Trp含量过高或过低的饲粮,会破坏体内的氨基酸平衡,从而导致氮的利用率降低,进而对鹅的生长性能产生负面影响,而适当含量的Trp则能够维持氨基酸的平衡,调节机体的蛋白质代谢,使得机体蛋白质沉积增加,从而改善动物的生长性能。
3.3 色氨酸对扬州鹅屠宰性能的影响 目前国内外有关Trp对屠宰性能的影响主要集中在肉鸡、蛋鸡上,而在水禽鹅上的研究较少。在肉鸡上的大量研究表明,饲粮中缺乏Trp时,肉鸡的屠宰率、胸肌率和腹脂率较低,补充晶体Trp后,动物的胸肌率和屠宰率显著提高,腹脂率呈现不同的变化规律 (周斌等,2011;席鹏彬等,2009;Corzo等,2005a)。本试验研究发现,Trp对扬州鹅屠宰率、全净膛率、半净膛率、腿肌率和腹脂率均无显著影响,而对胸肌率影响显著。其中,中Trp组胸肌率较低Trp组显著增加,高Trp组较低Trp组有增加的趋势。由此可见,饲粮Trp水平对于扬州鹅胸肌率的影响与对肉鸡胸肌率的影响基本上一致,而对于屠宰率和腹脂沉积的影响效果却不同。造成动物腹脂沉积增加的原因可能是由于动物采食较高Trp水平的饲粮,并且采食量提高,增加畜禽腹脂沉积,而有的腹脂沉积呈现下降趋势,这可能是由于不同品种脂肪的沉积规律不同而产生的。
4 小结
4.1 饲粮中添加适当水平的Trp能够提高5~10周龄扬州鹅平均日采食量和平均日增重,Trp水平过低或过高均会对扬州鹅生长性能产生不利影响。
4.2 随着饲粮中Trp水平的增加,5~10周龄扬州鹅胸肌率显著增加,而不同Trp水平则对屠体率、全净膛率,半净膛率、腿肌率和腹脂率均无显著影响。
4.3 动物生长性能提高除了与其采食量增加有关外,Trp在改善生长性能的过程中也发挥了重要的作用。
[1]饶巍,王玥,周斌,等.日粮色氨酸水平对蛋鸡生产性能及蛋白质代谢的影响[J].中国畜牧杂志,2011,47(15):38 ~ 41.
[2]王勇生,侯水生,刘福柱,等.0~21日龄北京鸭色氨酸、苏氨酸、异亮氨酸需要量的研究[J].西北农林科技大学学报(自然科学版),2005,33(7):25 ~ 30.
[3]席鹏彬,林映才,蒋宗勇,等.饲粮色氨酸对43~63日龄黄羽肉鸡生长、胴体品质、体成分沉积及下丘脑5-羟色胺的影响 [J].动物营养学报,2009,21(2):137 ~ 145.
[4]杨宁.家禽生产学[M].北京:中国农业出版社,2002.
[5]张括,王安,刘洋景.12~17周龄金定蛋鸭色氨酸适宜需要量的研究[J].饲料工业,2011,32(20):6 ~ 9.
[6]周斌,李慧,邹晓庭,等.色氨酸对产蛋鸡脂肪代谢的影响[J].中国畜牧杂志,2011,47(7):50 ~ 53.
[7]Adeola O,Ball R O.Hypothalamic neurotransmitter concentrations and meat quality in stressed pigs offered excess dietary tryptophan and tyrosine[J].J Anim Sci,1992,70(6):1888 ~ 1894.
[8]Adeola O,Ball R O.Hypothalamic neurotransmitter concentrations and meat quality in stressed pigs offered excess dietary tryptophan and tyrosine[J].J Anim Sci,1992,70(6):1888 ~ 1894.
[9]Andrew A F,Frank H,Markus R.Estimates of individual factors of the tryptophan requirements based on protein and tryptophan accretion responses to increasing tryptophan supply in broiler chickens 8~21 days of age[J].Arch Anim Nutr,2005,59(3):181 ~ 190.
[10]Corzo A,Kidd M T,Thaxton J P,et al.Dietary tryptophan effects on growth and stress responses of male broiler chicks[J].Brit Poultry Sci,2005b,46(4):478 ~ 484.
[11]Corzo A,Moran E T,Hoehler Jr D,et al.Dietary tryptophan need of broiler males from forty-two to fifty-six days of age[J].Poultry Sci,2005a,84(2):226 ~ 231.
[12]Denbow D M,Hobbs F C,Hulet R M,et al.Supplemental dietary L-tryptophan effects on growth,meat quality,and brain catecholamine and indoleamine concentrations in turkeys[J].Brit Poultry Sci,1993,34(4):715 ~ 724.
[13]Emadi M,Jahanshiri F,Kaveh K,et al.Tryptophan stimulates immune response in broiler chickens challenged with infectious bursal disease vaccine[J].J Anim Vet Adv,2010,9(3):610 ~ 616.
[14]Fatufe A A,Hirche F,Rodehutscord M.Estimates of individual factors of the tryptophan requirement based on protein and tryptophan accretion responses to increasing tryptophan supply in broiler chickens 8~21 days of age[J].Arch Anim Nutr,2005,59(3):181 ~ 190.
[15]Harms R H,Russell G B.Evaluation of tryptophan requirement of the commercial layer by using a corn-soybean meal basal diet[J].Poultry Sci,2000,79(5):740 ~ 742.
[16]Page T G,Southern L L,Ward T L.Effect of dietary tryptophan in excess of the requirement on growth and carcass characteristics of finishing pigs[J].Pro Anim Sci,1993,9:86 ~ 88.
[17]Rosa A P,Pesti G M,Edwards H M Jr,et al.Tryptophan requirements of different broiler genotypes[J].Poultry Sci,2001,80(12):1718 ~ 1722.
[18]Rosebrough R W.Crude protein and supplemental dietary tryptophan effects on growth and tissue neurotransmitter levels in the broiler[J].Brit J Nutr,1996,76(1):87 ~ 96.
[19]Shan A S,Sterling K G,Pesti G M,et al.The influence of temperature on the threonine and tryptophan requirements of young broiler chicks[J].Poultry Sci,2003,82(7):1154 ~ 1162.
