对牙鲆进行一代选择之后的育种效果分析
2012-10-08刘旭东刘志鹏王亚楠刘金相王志刚刘寿堂张全启
刘旭东,刘志鹏,王亚楠,刘金相,王志刚,刘寿堂,张全启
(1.中国海洋大学 海洋生命学院海洋生物遗传学与育种教育部重点实验室,山东 青岛,266003;2.国家海洋局 北海环境监测中心,山东 青岛 266033;3.海阳市黄海水产有限公司,山东 海阳,265122)
牙鲆(Paralichthysolivaceus)属于鲽形目牙鲆科牙鲆属,是我国重要的海水养殖鱼类品种之一,主要分布在我国渤海、黄海、南海以及朝鲜半岛、日本、俄罗斯远东沿岸海区,具有较高的经济价值,在渔业生产中占有重要的地位。我国的牙鲆增养殖业从20世纪80年代开始得到了迅猛发展,苗种需求量逐年增大,但苗种培育主要依赖于捕捞野生亲鱼和人工累代繁殖的亲本。牙鲆的高繁殖能力使得国内大部分养殖基地亲本繁育群体过小和更换率过低,加之亲本对后代的遗传贡献率存在差异,造成近交积累,导致牙鲆养殖群体遗传多样性降低[1-2]。同时伴随着养殖环境的恶化,养殖的牙鲆疾病爆发频繁、死亡严重、生长速度减慢、养殖效益大幅下降。因此有必要利用现有的不同背景的牙鲆亲鱼群体,依据数量遗传学原理,使用选择育种的方法,针对某些重要经济性状进行遗传改良,培育出高产、抗逆的新品种,从而奠定牙鲆养殖业健康高效的发展基础。
20世纪70年代以来,国外研究人员在水产动物的研究上,陆续开展了虹鳟(Salmogairdneri)[3]、马氏珠母贝(Pinctadafucatamartensi)[4]、海湾 扇贝(Argopectenirradians)[5]、罗非 鱼 (Oreochromisniloticus)[6]、大西洋鲑(Salmosalar)[7]等的选择育种,并且取得令人瞩目的进展。我国在水产动物选择育种方面,也相继开展了罗氏沼虾[8]、马粪海胆[9]、刺参耳状幼体[10]、大菱鲆[11]、中国对虾[12-13]、牙鲆[14]、哲罗鲑幼鱼[15]等水产动物的选择育种研究。
在选择育种中,遗传参数的准确估计是一项基础工作,它不仅是基因型选种的基础,而且也是制定育种规划和个体遗传评定的重要参数[16]。由于遗传力是性状、群体和环境三者特性的综合体现,因此,群体遗传结构和环境条件的改变,都会影响到性状遗传力。如果对任何情况均采用同一不变的遗传参数,则选择的实际效果与期望的进展必然存在较大的偏差。动物选择育种工作的主要目的是提高重要经济性状的生产性能,取得最大的经济效益。因此,育种工作的有效性主要是以选择所取得的遗传进展来衡量的,而遗传进展的高低主要取决于育种所用亲代的生产性状的遗传力的大小。对低等遗传力的个体或群体进行选育,后代对选择的反应较小,只有对中等以上遗传力的个体或群体进行选择,才会获得较理想的选择效果。对养殖物种的遗传改良,往往需要进行连续多代选择育种,而经过每一代选择后,都需要对上一代的选择效果进行评估和确认。同时,也需要对选育的下一代的生产性状遗传力进行精确评估,以确定进一步选择的有效性,制定正确的育种方案。在牙鲆选择育种方面,严复升等[14]完成了3月龄牙鲆体质量的主要形态性状的通经分析,评估体重与形态形状的相关关系,为牙鲆选育提供数据支持和测度指标,但未对选育群体的遗传力进行评估。刘永新和刘海金用不同模型估计牙鲆第一代家系间生长性状遗传参数,但未进行累代选育,因此也没有对选育一代后的遗传力进行评估[17]。
通过选择育种,我们已经培育出了牙鲆的子二代群体。基于牙鲆具有高繁殖性能的特性,依据巢式交配设计的方法,以选育的第一代个体为亲本,建立了第二代全同胞和半同胞家系,通过收集资料,以牙鲆第二代子代测定的体重、体高、全长为指标对第一代选育的效果进行评估,同时对牙鲆第二代家系群体的全长、体高和体重等生长性状的遗传力进行了估计,旨在为牙鲆连续多代选择育种提供宝贵的原始参数材料,为建立牙鲆进一步的育种策略、制定正确可行的育种方案提供必要的理论依据。
1 材料与方法
1.1 牙鲆家系的建立与养殖
以2010年建立的牙鲆家系为试验材料,家系建立于2010-05中旬,试验用亲鱼为经过家系选育的具有优良生长性状的第一代个体。依据个体性腺发育成熟程度、无病症和外伤等情况选择亲鱼,将没有亲缘关系的个体进行配组,以雌、雄比例为2∶1,建立半同胞家系;以雌、雄比例为1∶1,建立全同胞家系。共建立10个父系半同胞家系及20个全同胞家系。同时,建立以未经选择的野生牙鲆为亲本的混合群体作为对照群体,各个家系及混合群体均培养在直径1.5m、深1.3m的培养缸内,所有家系的培养条件尽量保持一致:水深1.0m、温度(18±1)℃、连续充气、每天换水30%,4月龄时各家系随机取24~50尾牙鲆,记录体重、体高和全长。
1.2 数据分析
差异性分析用SPSS13.0软件的单因素方差分析检验法,事先均进行并通过正态性检验和方差齐性检验。生长性状数据的方差分析使用SPSS软件的一般线性模型(General line model,GLM)。二因素系统分组的方差分析的数学模型为:xijk=u+si+dij+eijk,其中,xijk表示某性状第k个个体的生长指标,u表示总体均值,si表示第i个雄性效应,dij表示第i个雄性内第j个个体的雌性效应,eijk表示随机误差变量。全同胞资料表型变量的方差组成如表1所示。

表1 表型变量组成的方差分析Table 1 Variance analysis of phenotypic variation
其中k1为牙鲆雄性内雌性间雌性个体的有效平均后代数目,k2为雌性个体的有效平均后代数目,k3为雄性个体的有效平均后代数目,计算公式:
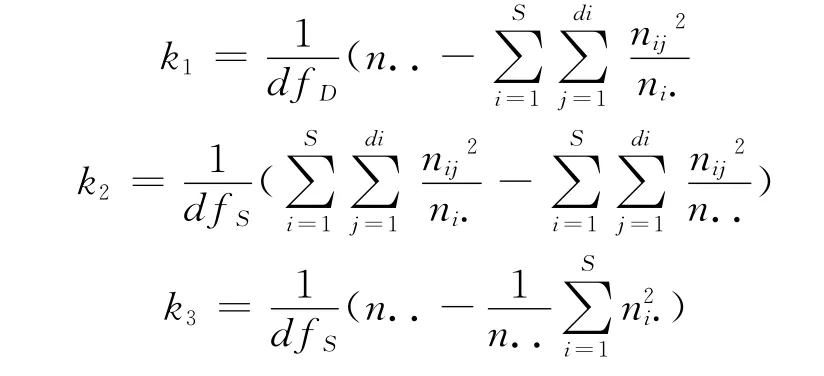
式中,n..为全部后代数总和;ni·为第i尾雄亲与配对的所有雌亲的后代个体总数;nij为第i尾雄亲与配对的第j尾雌亲的后代个体总数。
全同胞资料估计的遗传力:分别为父系半同胞hS2=4×σS2/(σS2+σD2+σe2);母系半同胞hD2=4×σD2/(σS2+σD2+σe2);全同胞估计的狭义遗传力为全同胞组内相关系数的2倍,即hSD2=2×(σS2+σD2)/(σS2+σD2+σe2)。
2 结 果
2.1 牙鲆选育第二代的经济性状遗传进展
经过选择的牙鲆各家系留种率为0.20,利用的对照群体来源于未经选育的牙鲆群体混合子代,群体实际测量结果见表2,群体间差异结果见表3。

表2 选育第二代牙鲆4月龄的生长参数测量Table 2 Growth Parameters of the second generation of P.olivaceus at 4months
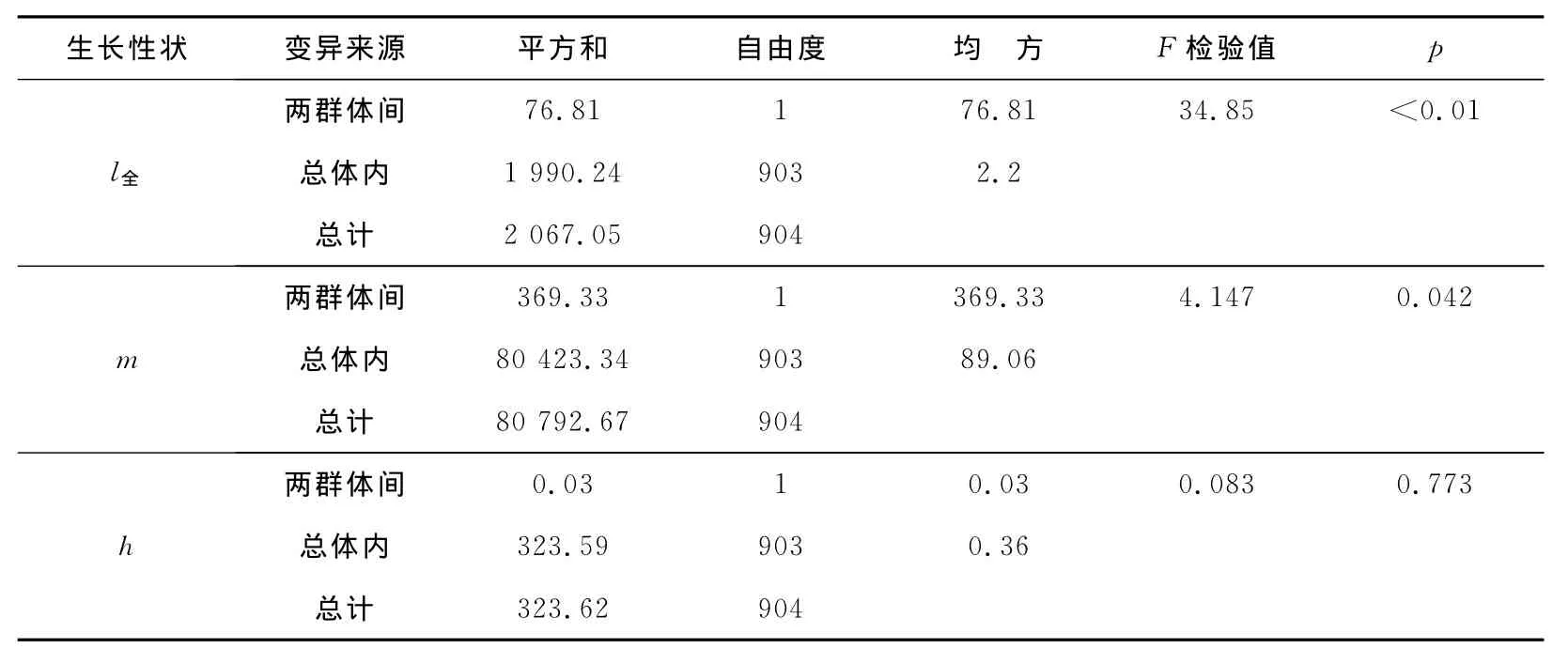
表3 选育群体和对照群体生长参数差异的方差分析Table 3 ANOVA for growth parameters between selected group and control group
结果表明,选育第二代群体的平均体质量比未经选育的对照提高2.91g,遗传增益为11.25%,显著高于未经选育的群体(p<0.05);平均全长较未选育的对照提高1.33cm,遗传增益为10.36%,极显著高于未经选育的群体(p<0.01);选育群体和对照群体的体高无显著差异。
2.2 牙鲆体重体长体高遗传力估计及显著性检验
通过SPSS软件的分析,得到牙鲆体重、全长、体高资料表型变量的方差组成和分析见表4及表5。通过计算牙鲆雄性内雌性间雌性个体的有效平均后代数目(k1)、雌性个体的有效平均后代数目(k2)以及雄性个体的有效平均后代数目(k3),结果为k1=42.83,k2=42.95,k3=85.65。最终计算得到体重、体长,体高遗传力分别为0.656~0.731,0.481~0.742,0.480~0.651,显著性检验结果(表6)显示,父系半同胞方差组分估计的体重、体长、体高的遗传力均未达到显著水平(t=1.06~1.36),母系半同胞方差组分估计遗传力均达到显著水平(t=2.03~2.05),而全同胞组分方差组分的体重、体长,体高遗传力估计均达到极显著水平(t=2.58~2.69)。

表4 4月龄牙鲆表型变量组成的方差分析Table 4 Analysis of variance components of phenotypic variation of P.olivaceus at age of 4months

表5 表型变量的原因方差组分Table 5 Causal variance components of phenotypic variances

表6 各性状遗传力估计及t检验Table 6 Estimation of heritability for different growth-related traits and t-test
3 讨 论
3.1 选择育种
选择育种是根据育种目标,在现有育种材料内将出现自然变异的类型,经比较鉴定,通过多种选择方法,选优去劣,选出优良的变异个体,培育新品种的方法[18]。其基本前提是利用选育群体存在较高的遗传变异,提高选择强度,随之可以提高育种后代群体相对与人类有益的优良基因型比率,得到理想的遗传改进结果。选择育种具有长期性,一个世代的选择,得到的遗传改良是有限的,只有通过长期的选择和定向的培育,才能使目标性状朝着期望的方向发展并一步一步的固定。Haskin和Ford对美洲牡蛎(Crassostreavirginica)进行连续4代的抗单胞子虫病的选择,使其抗病能力比野生群体提高了8~9倍[19]。日本学者对真鲷生长速度进行了连续6代选择育种,从第三代开始,选育群体表现出明显的生长优势,经过6代选育后,生长速度比选育前明显提高,商业化养殖周期由原来的3~4a缩短到2a[20],挪威AKVAFORSK遗传研究所自从20世纪70年代以来运用BLUP选择育种,使大西洋鲑的7个性状已经得到改良,目前选育的性状达到10个以上,他们选育的大西洋鲑已被广泛地应用于生产养殖,使挪威大西洋鲑的养殖成为世界上最成功的海水养殖业[21]。李健等经过连续6代的群体选育,培育出表现出生长快、抗逆性强等优良的经济性状中国对虾快速生长新品种“黄海1号”[12]。孔杰等1998-04开始了中国对虾抗病良种的选育,采用群体、家系与多性状BLUP复合育种技术,经过10代连续选育获得了第1个具有多个优良性状(生长快、抗wssv性能强、养殖存活率高)的“黄海2号”对虾系种[13]。
方差分析所选择改良的群体表明,初步达到改良生长性状的效果。经过选育的群体体质量显著高于未经选育的群体(p<0.05),且体质量的提高了2.91g,遗传增益为11.25%;经过选育的群体全长极显著高于未经选育的群体(p<0.01),且全长的遗传增益较未选育的群体提高了1.33cm,遗传增益为10.36%。而体高的差异不显著,分析原因可能是因为体高与体重、体长的回归系数较高,遗传增益反应到体高上时效应不明显;同时由于体高的标准差很小,人为测量的误差因素也有可能造成此种结果。
研究中同时发现在雄亲间和雌亲间遗传一致性较低,各表型性状间均存在极显著差异(表4),具有进一步的选择和改良的潜力。
3.2 遗传力的估计
任何一个群体经过选择之后,群体的基因频率发生变化,群体的性状遗传力是随着不同选择方法和目标基因纯合程度而变化的。如果对任何情况均采用同一不变的遗传参数,则选择的实际效果与期望的进展必然存在较大的偏差。因此,在经过一代或多代选择之后,对选择的下一代进行性状的遗传力估计,是制定进一步选择计划的理论依据。我们曾对牙鲆进行了一代家系选育,在本研究中通过选育后代的遗传进展对第一代选育效果进行评估,同时对经过一代选育后的第二代牙鲆群体进行生长性状的遗传力估计,依次为今后选育提供育种设计的理论参考。刘永新和刘海金[17]对240日龄牙鲆体重、体长、体高的遗传力的估计值分别为0.30,0.32,0.39。严复升等[14]完成了3月龄牙鲆体质量的主要形态性状的通经分析,评估体重与形态形状的相关关系。我们分析得出牙鲆第二代体重、全长和体高的遗传力分别为0.656~0.731,0.481~0.742,0.480~0.651,高于前两者的研究结果。其原因可能是由于遗传力估计所用的样本的年龄造成的,不同生长阶段的牙鲆其生长速度差异不同,测算的遗传力也有差距。另外,可能是因为不同群体遗传结构本身就不同,其培育的环境、年份等也有差别,例如育种开始时间、养殖水温、日换水量、养殖密度、饲料投喂因素、数据测量时间等,这些变化均可造成遗传力估计结果差异。
在各遗传力的所有估计值中,父系半同胞、母系半同胞和全同胞个体间遗传力的估计值是有差别的,经统计t检验,母系半同胞个体间遗传力的估计值达到显著水平,全同胞个体间遗传力的估计值达到极显著水平,其中体高和体长的母系半同胞遗传力估计值最高,表明存在着较大的母性效应。这可能是由于卵细胞质量、上位效应和环境效应共同作用的结果。预示对母本的的选择效果可能优于对本的选择。
Maluwa对罗非鱼体重的选择响应进行了研究,其F1,F2,F3代的遗传力分别为(0.31±0.09),(0.31±0.10),(0.35±0.1)[22]。在鲤鱼研究中,其13个月的体长和体重遗传力分别为0.50和0.5[23]。经过7代选择育种所获得的中国对虾新品种“黄海1号”3月龄和4月龄体长的遗传力估计值分别在0.46~0.53和0.44~0.48[24]。由此看出,我们所选育的第二代牙鲆群体的体重、全长和体高的遗传力都为中等或偏高遗传力。这些性状仍然具有较大的选择育种潜力,在今后的牙鲆选择育种中仍旧可以取得较快的遗传进展。我们认为有必要在今后的选育过程中,不断重复测量不同时期、不同环境条件下的数据,得到更为准确的生产性状遗传参数,借助数量遗传学方法制定更有效的育种规划,对牙的生长性状进行选择,更快更好地培育牙鲆养殖新品种。
(References):
[1] ZHANG Q Q,XU X F,QI J,et al.The genetic diversity of wild and farmed Japanese Flounder populations[J].Periodical of Ocean University of China,2004,34(5):816-820.张全启,徐晓斐,齐洁,等.牙鲆野生群体与养殖群体的遗传多样性分析[J].中国海洋大学学报,2004,34 (5):816-820.
[2] WANG W,YOU F,GAO T X,et al.Genetic variations at ten microsatellite loci in natural and cultured stocks of left-eyed flounderParalichthysolivaceusin Shandong coastal waters[J].Oceanologia et Limnologia Sinica,2004,35(6):530-538.王伟,尤锋,高天翔,等.山东近海牙鲆自然和养殖群体10个微卫星基因座位的遗传多态性分析[J].海洋与湖沼,2004,35(6):530-538.
[3] AULSTAD D,GJEDREM T.Genetic and environmental sources of variation in length and weight of rainbow trout(S.gairdneri)[J].Fisheries Research Board of Canada,1972,29:237-341.
[4] WADA K.Genetic selection for shell traits in the Japanese pear oyster,Pinctadafucatamartensi[J].Aquaculture,1986,57:171-176.
[5] GRENSHAW J,HEFFERNAN P,WALKER R.Heritability of growth rate in the southern Bay scallop,Argopectenirradiansconcentricus[J].Journal of Shellfish Research,1991,10:55-63.
[6] GRADAM A,BAKAR Y.Application of mixed-model techniques to fish breed improvement:analysis of breeding-value selection to increase 98-day body weight in tilapia[J].Aquaculture,2002,212:93-113.
[7] QUINTON C,MCMILLAN I,GLEBE B.Development of an Atlantic salmon(Salmosalar)genetic improvement program:genetic parameters of harvest body weight and carcass quality traits estimated with animal models[J].Aquaculture,2005,247:211-217.
[8] CHEN G,CAI H J,LIN X W.Primary analysis in genetic parameters ofMacrobrachiumrosenbergii[J].Journal of Zhanjiang Fisheries College,1996,16(1):25-30.陈刚,蔡华紧,林晓文.罗氏沼虾体长和体重的一些遗传参数分析[J].湛江水产学院学报,1996,16(1):25-30.
[9] LIU X L,CHANG Y Q,XIANG J H,et al.Heritability of juvenile growth for the sea urchinsStrongylocentrotusintermedius[J].Journal of Fishery Sciences of China,2003,10(3):206-211.刘小林,常亚青,相建海,等.虾夷马粪海胆早期生长发育的遗传力估计[J].中国水产科学,2003,10(3):206-211.
[10] LUAN S,SUN H L,KONG J.Heritability of auricularia larval body length for sea cucumberApostichopusjaponicusSelenka[J].Journal of Fishery Sciences of China,2006,13(3):378-383.栾生,孙慧玲,孔杰.刺参耳状幼体遗传力的估计[J].中国水产科学,2006,13(3):378-383.
[11] MA A J,WANG X A,YANG Z,et al.The growth traits and their heritability of young turbot(ScophthalmusmaximusL.)[J].Oceanologia et Limnologia Sinica,2008,39(5):499-504.马爱军,王新安,杨志,等.大菱鲆(ScophthalmusmaximusL.)幼鱼生长性状的遗传力及其相关性分析[J].海洋与湖沼,2008,39(5):499-504.
[12] LI J,LIU P,HE Y Y,et al.Artificial selection in the new breed ofFenneropenaeuschinensisnamed“Yellow Sea 1”based on fast growth trait[J].Journal of Fisheries of China,2005,29(1):1-5.李健,刘萍,何玉英,等.中国对虾快速生长新品种“黄海1号”的人工选育[J].水产学报,2005,29(1):1-5.
[13] MENG X H.The Analysis of WSSV resistance ofFenneropenaeuschinensis“Huanghai No.2”[D].Qingdao:Ocean University of Chi-na,2010.孟宪红.中国对虾“黄海2号”对WSSV的抗病性分析[D].青岛:中国海洋大学,2010.
[14] YAN F S,WANG Z G,LIU X D,et al.Path analysis of the effects of morphometric traits on body weight for 3-month agedParalichthysolivaceus[J].Progress In Fishery Sciences.2010,31(2):45-50.严福升,王志刚,刘旭东,等.3月龄牙鲆形态性状对体质量的通径分析[J].渔业科学进展,2010,31(2):45-50.
[15] WANG J,KUANG Y Y,TONG G X,et al.Genetic parameters of growth traits inHuchotaimenat different temperature[J].Journal of Fishery Sciences of China,2011,18(1):75-82.王俊,匡友谊,佟广香,等.不同温度下哲罗鲑幼鱼生长性状的遗传参数估计[J].中国水产科学,2011,18(1):75-82.
[16] ZHANG Y,ZHANG Q.Linear model in livestock breeding[M].Beijing:Beijing Agricultural University Press,1993:52-86.张沅,张勤.畜禽育种中的线性模型[M].北京:北京农业大学出版社,1993:52-86.
[17] LIU Y X,LIU H J.Comparison and analysis of genetic paramaters of growth traits estimatied by different models for Japanese flounder families[J].Progress in Fishery Sciences,2010,31(1):40-47.刘永新,刘海金.不同模型估计牙鲆家系间生长性状遗传参数的比较分析[J].渔业科学进展,2010,31(1):40-47.
[18] FAN Z T.Aquatic animal genetics and breeding[M].Beijing:China Agricultural Press.2005:58-68.范兆廷.水产动物育种学[M].北京:中国农业出版社,2005:58-68.
[19] HASKIN L,FORD S.Development of resistance toMinchinianelsoni(MSX)mortality in laboratory reared and native oyster stocks in Delaware Bay[J].Marine Fisheries Review,1979,41:54-59.
[20] MURATA O,HARADA S,MIYASITA K,et al.Selective breeding for growth in red sea bream[J].Fisheries Science,1996,62(6):845-849.
[21] LUAN S,KONG J,WANG Q Y.Methods and application of aquatic animal breeding value estimation:A review[J].Marine Fisheries Research,2008,29(3):101-107.栾生,孔杰,王清印.水产动物育种值估计方法及其应用的研究进展[J].海洋水产研究,2008,29(3):101-107.
[22] MALUWA.Response to selection for harvest body weight ofOreochromisshiranus[J].Aquaculture,2007,273:33-44.
[23] BONGERS A,BOVENHUIS H,VAN STOKKOM A,et al.Distribution of genetic variance in gynogenetic or androgenetic families[J].Aquaculture,1997,153:225-238.
[24] HUANG F Y,HE Y Y,LI J,et al.Estimates for the heritability of body length in shrimpFenneropenaeuschinensisNamed“Huanghai No.1”[J].Periodical of Ocean University of China,2008,38(2):269-274.黄付友,何玉英,李健,等.“黄海1号”中国对虾体长遗传力的估计[J].中国海洋大学学报,2008,38(2):269-274.
