酿酒酵母中线粒体活性对甲酸处理下细胞死亡率的影响*
2012-09-12杜林李莲
杜林,李莲
1(仲恺农业工程学院轻工食品学院,广东广州,510225)
2(中山大学生命科学院,广东广州,510275)
酿酒酵母中线粒体活性对甲酸处理下细胞死亡率的影响*
杜林1,李莲2
1(仲恺农业工程学院轻工食品学院,广东广州,510225)
2(中山大学生命科学院,广东广州,510275)
已知甲酸可抑制线粒体呼吸链末端的细胞色素氧化酶,低浓度的甲酸可导致细胞内线粒体活性氧的快速爆发并进一步杀死酵母细胞。因此,对可能影响酿酒酵母细胞线粒体产生活性氧的因素进行了研究,分析了菌龄、低温预处理、还原剂、线粒体ATP酶基因突变等对甲酸毒性的影响。结果表明,酿酒酵母对甲酸的抗性与线粒体活性密切相关,通过线粒体突变降低线粒体活性可增强酿酒酵母对甲酸的抗性。
酿酒酵母,甲酸抗性,线粒体,活性氧
甲酸是结构最简单的有机酸,也是一种弱有机酸。甲酸为酵母菌等微生物的代谢产物,但在浓度较高时其对酵母菌本身具有毒性。以纤维素为原料生产酒精是目前生物能源研究领域的一个重要内容。纤维素可以用理化方法和生物方法进行降解并以酿酒酵母发酵,但酿酒酵母在纤维素的水解液中的生长情况往往并不理想[1-2]。甲酸由于生成的浓度较高,而最低抑菌浓度比乙酸等其他弱有机酸低十倍以上,是纤维素水解液中主要的抑菌因素之一。因此,酵母菌的甲酸抗性突变株在纤维素发酵产酒精中具有较大的价值。另外,在微生物制取氢气方面,一个很有吸引力的方法是以酿酒酵母转化各种生物原料产生甲酸,再以大肠杆菌等的氢化酶将甲酸分解为CO2和H2。但酿酒酵母难以耐受20 mmol/L以上的甲酸,因此获得甲酸抗性突变株可以大大提高甲酸的积累浓度,并促进这一技术的应用[3]。但是,由于对甲酸细胞毒性机理的认识不充分,目前并没有见到构建强甲酸抗性酵母菌株的报道。
甲醇在人体肝脏内会被代谢为甲醛,并进一步氧化为甲酸。因此在医学领域甲酸的毒性机理已得到较多研究[4]。已经有研究者证明,甲酸和CO、氰化物、AS2O3等具强细胞毒性的化合物类似,可以和线粒体内膜上的细胞色素氧化酶上亚铁离子发生结合,抑制细胞色素氧化酶的活性[5-8]。我们据此推测,甲酸导致细胞快速死亡的机理不是由于细胞内能量的耗尽,而是因为它引起细胞色素氧化酶活性的异常,导致活性氧的产生、爆发,进而氧化破坏细胞。我们的初步研究表明,线粒体产生的活性氧和甲酸诱导的酵母细胞死亡关系密切[9]。因此,我们推测,降低酵母菌线粒体活性,抑制活性氧的产生,可以增强酿酒酵母对甲酸的抗性。
1 材料与方法
1.1 仪器与设备
恒温摇床(哈尔滨东明仪器HZQ-C);培养箱(Thermo);流式细胞仪(Vantage SE,BD Biosciences)。
2.2 酵母菌菌株、质粒及培养基
酿酒酵母BY4741由美国Rochester大学的David Goldfarb教授惠赠。酿酒酵母W303-1a野生型菌株由中山大学微生物遗传实验室提供,W303-1a背景ATP酶4亚基基因缺失株由中山大学微生物遗传实验室构建。
YPD液体培养基:2%蛋白胨,1%酵母提取物,2%D-(+)-葡萄糖,分装后121℃高压灭菌20 min。
2.3 酵母细胞的培养及甲酸处理后的酵母细胞存活率测定
取对数期酿酒酵母细胞培养液0.5 mL,接种至装于250 mL三角瓶中的50 mL YPD液体培养基中,置于30℃,200 r/min往复式振荡培养箱培养细胞至对数期或稳定期,分别取1 mL菌液于1mL离心管中,立即加入相应摩尔浓度的甲酸,30℃处理40 min后,在YPD培养基上以标准平板计数法计数,每一稀释度涂3个平板,以3次独立的实验结果计算存活率。
2.6 细胞内活性氧爆发的流式细胞仪检测
细胞质中活性氧检测:取5 mL YPD加入试管中,接入50 μL过夜培养的酵母菌液,摇床培养6h(30℃,200 r/min)。吸取以DMSO为溶剂的10 mmol/L双乙酸二氯荧光黄(dichlorofluorescein diacetate,DCFH-DA)溶液2 μmol/L,使染色时DCFH-DA的工作浓度为20 μmol/L,避光孵育15 min,使DCFH-DA充分进入细胞。根据需要,迅速加入一定量的1mol/L甲酸,室温或培养箱恒温避光处理,根据实验要求,分别吸取100 μL处理中的各样品菌液加入FACS管中,加入900 μL冷藏的PBS,立即以流式细胞仪上样分析。所用氩离子激光器激光波长为488 nm,在530 nm检测激发出的荧光,以流式细胞仪FL-1通道进行检测。或加入PI后以FL-1和FL-2通道检测。
超氧阴离子的检测:先以无菌DMSO配置10 mmol/L的超氧化物阴离子荧光探针二氢溴化乙锭(Dihydroethidium,DHE)溶液,取0.5 μL加于1 mL菌液,使DHE的终浓度为5 μmmol/L,30℃避光孵育15 min,加入相应浓度甲酸避光处理,以流式细胞仪FL-2通道进行分析。激发光波长为535 nm,发射光波长为610 nm。
3 结果与分析
3.1 甲酸处理导致线粒体超氧阴离子的产生与自由基的扩散
为了了解活性氧爆发时细胞内超氧阴离子及细胞质内过氧化氢等活性氧的水平,故使用活性氧荧光探针DCFH-DA和DHE同时对甲酸处理时细胞中的活性氧进行检测。

图1 FACS分析甲酸诱导的细胞质活性氧和超氧阴离子产生
由图1可见,在用甲酸处理10 min时,以荧光探针DCFH-DA和DHE依次检测到活性氧的产生。细胞内DHE阳性率超过DCFH-DA的阳性率,显示DHE氧化后的荧光先产生。这和两种荧光染料的特性有关,DHE可检测线粒体产生的超氧阴离子,DCFH-DA可反映细胞质中的活性氧水平。线粒体电子传递链障碍导致活性氧产生时,超氧阴离子首先产生。所以实验中首先检测到较多的细胞产生了超氧阴离子,而此时以DCFH-DA检测到的细胞质活性氧阳性的细胞却比超氧阴离子阳性的细胞少,说明细胞质内的活性氧是由线粒体内超氧阴离子扩散形成。
3.2 还原剂预处理对活性氧爆发及存活率的影响
N-乙酰半胱氨酸(N-acetylcysteine,NAC),是细胞内还原性谷胱甘肽的前体,因为能够快速穿过细胞膜,因此是一种细胞生物学研究中广泛使用的还原剂。NAC可在细胞内形成含巯基的代谢产物,促进GSH的合成,消除自由基。在用甲酸处理酵母细胞,使其细胞内活性氧爆发前我们先用NAC预处理细胞,使其细胞内NAC达到较高浓度以抑制活性氧的水平,检测抑制活性氧对细胞存活率的影响。

图2 还原剂NAC预处理对60 mmol/L甲酸处理后死亡率的影响
由图2可见,和对照相比,2 mmol/L的NAC预处理明显降低了死亡率,说明细胞内还原剂的增加,活性氧的抑制可降低甲酸处理后细胞的死亡率。有可能是由于NAC减少了活性氧,从而抑制了活性氧对酵母细胞的损伤。另外,使用PI染色后流式细胞仪分析细胞膜完整性的方法也证明NAC预处理可以降低甲酸处理后的酵母细胞死亡率。
3.3 酿酒酵母线粒体活性对甲酸处理下细胞存活率的影响
在病理和生理条件下,通常线粒体都是活性氧的主要来源。线粒体电子传递链的障碍,往往导致活性氧的大量形成。而在不同遗传性状的酿酒酵母细胞中,或同一种细胞处在不同的生长条件下,其线粒体活性往往有很大差异[10]。因此我们对不同类型酿酒酵母细胞中活性氧的爆发及细胞死亡情况进行了研究。
3.3.1 酵母细胞菌龄对甲酸处理时细胞死亡率的影响
和稳定期细胞相比,对数期的酵母细胞代谢更为旺盛,其线粒体活性更高,因此我们推测同浓度甲酸在对数期酵母细胞中能导致更猛烈的活性氧爆发,从而对细胞造成更大的破坏,因此用甲酸处理培养24 h的稳定期细胞时活性氧产生情况,并比较培养6 h的对数期细胞和培养24 h的稳定期细胞在同浓度甲酸处理时的存活率。
由图3可见,对数期细胞对甲酸的敏感程度比稳定期细胞高得多,这在甲酸浓度较高时更为明显。故甲酸的毒性和酵母细胞的代谢状态密切相关。因此可以认为,对数期细胞由于处于代谢旺盛的特点,其细胞内较高活性的线粒体在甲酸处理下产生更多的活性氧,从而导致更多的对数期细胞死亡。
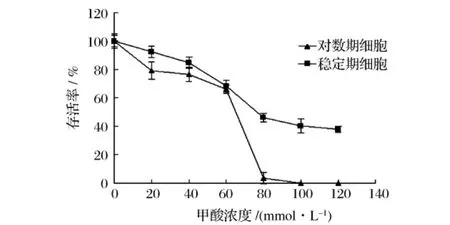
图3 甲酸处理后对数期及稳定期细胞的存活率比较
3.3.2 低温预处理对甲酸诱导的细胞死亡率的影响
由于线粒体的产能速度和微生物细胞外环境密切相关,即细胞外因素会影响线粒体活性,因此我们推测低温预处理降低线粒体活性后,再以甲酸处理细胞,这时线粒体活性氧爆发应该减弱,细胞的死亡率应该下降。故在加入甲酸之前,先将细胞置于4℃10 min,再将其和正常培养的对照样品同时加入甲酸处理40 min,然后以标准平板计数法比较二者的存活率。在本实验中,我们还对另一实验室研究常用的酿酒酵母BY4741进行了测定。
由图4可见,短时间的低温预处理,即可明显提高甲酸处理后的细胞存活率。该结果也表明,甲酸诱导细胞死亡和细胞的代谢状态有关。

图44℃预处理对60 mmol/L甲酸诱导的细胞死亡率的影响
3.3.3 线粒体呼吸缺陷对甲酸处理细胞存活率的影响
ATP酶结构上的缺陷会影响其合成ATP的能力及线粒体的整体活性。所以我们通过敲除ATP酶亚基基因的方法,降低线粒体的活性,以期减弱甲酸诱导的活性氧的爆发并提高细胞存活率,并采用标准平板计数法对同浓度甲酸处理后野生型和ATP酶4亚基缺失株的存活率进行了比较。
由图5可见,线粒体缺陷会大大提高细胞的存活率,分析其原因,推测是在线粒体活性较低的情况下,甲酸诱导其产生的活性氧较少,故活性氧对细胞的破坏较小,所以细胞的存活率明显上升。此外,我们还发现位于线粒体外膜的AIF基因缺失后,其线粒体膜电位会下降,同时细胞对甲酸的抗性也会增强(数据未列出)。

图5 ATP酶亚基缺失对60 mmol/L甲酸处理细胞死亡率的影响
4 结论与讨论
活性氧是指一些能和还原性物质反应的含氧化合物。正常生理情况下,超氧阴离子的生成由NAD(P)H氧化酶、黄嘌呤氧化酶参与,也可由线粒体内电子传递链中的半醌类化合物参与[11],是线粒体内形成的第一种活性氧。由于甲酸可以阻断呼吸链,因此可以导致超氧阴离子的形成。超氧阴离子在线粒体内产生后,可以在线粒体内部或细胞质内转化成过氧化氢,因而在实验中可以用DCFH-DA检出较高比例的活性氧阳性细胞。在谷胱甘肽大量消耗的情况下,过氧化氢可破坏细胞成分,或进一步转化成氧化性更强的羟自由基,对核酸等结构产生致命的破坏。
正常细胞会产生少量的活性氧,而一些细胞强毒性试剂如抑制细胞色素氧化酶的砷化物、氰化物可导致活性氧的快速大量产生。本文研究表明,甲酸处理会使细胞迅速产生超氧阴离子,并进一步转化为细胞质中其他类型的活性氧。和稳定期的细胞相比,对数期酿酒酵母细胞对甲酸更为敏感,死亡率更高;还原剂NAC短时间预处理,较低温度预处理,敲除ATP酶的4亚基均可以降低细胞的死亡率。因此,降低甲酸处理下线粒体产生活性氧的能力可增强酿酒酵母细胞对甲酸的抗性。
因此,在深入研究甲酸毒性机理和抗性机制的基础上,利用分子生物学方法,改变酿酒酵母细胞内甲酸作用的靶位点或生化过程,构建线粒体呼吸链或其他线粒体膜蛋白的突变株,可获得具有较强甲酸抗性的菌株。
[1]Laluce C,Schenberg A C.Advances and Developments in Strategies to Improve Strains of Saccharomyces cerevisiae and Processes to Obtain the Lignocellulosic Ethanol-A Review[J].Appl Biochem Biotechnol,2012,166(8):1908-1926.
[2]Parawira W,Tekere M.Biotechnological strategies to overcome inhibitors in lignocellulose hydrolysates for ethanol production:review[J].Crit Rev Biotechnol,2011,31(1):20-31.
[3]Akinori Matsushika,Hiroyuki?Inoue,Ethanol production from xylose in engineered Saccharomyces cerevisiae strains:current state and perspectives[J].Applied Microbiology and Biotechnology,2009,84:37-53.
[4]陈捷敏,王立新,夏文涛.甲醇中毒机制的研究进展[J].法医学杂志,2010,26(4)294-296.
[5]Nicholls P.Formate as an inhibitor of cytochrome c oxidase[J].Biochem Biophys Res Commun,1975,67(2):610-616.
[6]Liesivuori J.Savolainen H.Methanol and formic acid toxicity:biochemical mechanisms[J].Pharmacol Toxicol,1991,69(3):157-163.
[7]Hantson P E.Acute methanol intoxication:physiopathology,prognosis and treatment[J].Bull Mem Acad R Med Belg,2005,160(5-6):294-300.
[8]Hantson P E.Acute methanol intoxication:physiopathology,prognosis and treatment[J].Bull Mem Acad R Med Belg,2006,161(6):425-434.
[9]Du Lin,Su Yingying,Sun Dongbei,et al.Formic acid induces Yca1p-independent apoptosis-like cell death in the yeast Saccharomyces cerevisiae[J].FEMS Yeast Research,2008(4):531-539.
[10]de Grey A D.Reactive oxygen species production in the mitochondrial matrix:implications for the mechanism of mitochondrial mutation accumulation[J].Rejuvenation Res,2005,8(1):13-17.
[11]Lenaz G.The production of reactive oxygen species:mechanisms and implications in human pathology[J].IUBMB Life,2001,52(3-5):159-164.
ABSTRACTBasing on the discovery that formic acid can inhibit the cytochrome oxidase which is located at the end of mitochondrial respiratory chain,and low concentration of formic acid can lead to the rapid outbreak of the reactive oxygen species which can kill yeast cells further,factors affecting the production of reactive oxygen species in Saccharomyces cerevisiae cells were studied.Effects of cell age,low-temperature,reducing agents pretreatment,and mitochondrial ATPase gene mutation to formic acid toxicity were analyzed.The results show that formic acid resistance of Saccharomyces cerevisiae is closely related to mitochondrial activity,reducing mitochondrial activity by mitochondrial mutation can enhance cellular resistance to formic acid.
Key wordsSaccharomyces cerevisiae,formic acid resistance,mitochondria,reactive oxygen species
Effect of Mitochondrial Activity to the Cell Survivla Rate of Saccharomyces cerevisiae After Formic Acid Treatment
Du Lin1,Li Lian2
1(Institute of Light Industry and Food Engineering,Zhongkai University of Agriculture and Engineering,Guangzhou 510025,China)
2(School of Life Science,Sun Yat-sen University,Guangzhou 510275,China)
教授(E-mail:dulin2000@163.com)。
*广东省产学研结合资助项目“广东客家黄酒的发酵菌种研究与工业化应用”。
2012-05-15,改回日期:2012-06-07
