法桐花粉主要过敏原基因Pla a1重组表达及鉴定①
2012-09-12王贵佐陶爱林孙秀珍李满祥邹泽红吴媛媛
王贵佐 陶爱林 孙秀珍 李满祥 刘 昀 邹泽红 吴媛媛
(西安交通大学医学院第二附属医院呼吸病研究室,西安710004)
法国梧桐为悬铃木科、悬铃木属,在各地广泛种植。Kosisky等[1]在1998~2007采用定量收集法对华盛顿市区进行了10年的气传花粉变应原调查,结果显示主要的花粉类型包括:栎树、柏树、松科、桑属、桦科、槭属、悬铃木属、梣属、禾本科,全年花粉总量中树花粉所占比例为91.2%,草类占7%。Aira等[2]对西班牙西北部Iberian Peninsula地区的主要绿化植物进行了调查,结果发现悬铃木属、齐墩果属是当地最主要的花粉来源。Ture等[3]报道了土耳其Bilecik地区气传花粉的研究,发现松属、柏科、悬铃木属、栎属、柳属是该地区的主要气传花粉,并且空气中花粉浓度最高在5月。Celenk等[4]在土耳其西北部的Bursa地区进行了气传花粉调查,发现松属、齐墩果属、悬铃木属、柏属、栎属、禾本科、荨麻科、栗属在空气花粉的播散中起主要作用,5月空气中花粉的浓度达到最高。法桐花粉不仅显示了较高的空气飘逸浓度,并且各地研究者发现法桐花粉可引起广泛的变态反应。刘光辉等[5]通过对湖北省气传花粉的调查并对2 300例花粉症患者进行皮肤过敏原检测试验及对发病季节进行分析,结果发现,湖北省阳性率最高的春季致敏花粉主要为悬铃木属,秋季致敏花粉主要为篙属、豚草属。卢家美等[6]通过对花粉的曝片采集及对168名过敏性哮喘患者进行花粉血液变应原筛查,得出艾蒿、法桐、藜草、葎草、禾本科可能是西安市的主要致敏花粉。Lauer等[7]发现在地中海地区法桐花粉变应原IgE反应率27.3% ~63.8%,是该地区的主要气传花粉变应原。越来越多的调查资料及临床流行病学资料表明,法国梧桐花粉是我国及西方多数国家地区花粉症的重要原因。
目前发现法桐变应原提取液中主要有以下主要致敏蛋白:Pla a1、Pla a2、Pla a3。Pla a1蛋白可与92%以上的法桐花粉症患者血清发生反应,而Pla a2、Pla a3 反应率分别为 84% 和 63.8%[7-9]。目前对法国梧桐花粉主要变应原的表达、纯化却鲜有研究,并且对Pla a1蛋白进行表达纯化具有很主要的现实意义。本研究对Pla a1蛋白首次进行了表达、纯化和鉴定,为制备高纯度变应原、重组低致敏过敏原及变应原核酸疫苗奠定基础。
1 材料与方法
1.1 试验材料 试验试剂:T4 DNA连接酶、pET-44a表达载体购自 Promega公司;宿主大肠杆菌JM109、表达宿主大肠杆菌Rosetta购自Novagen公司;DNA合成和测序由上海博尚生物公司完成;rTaq 聚合酶、10 ×PCR buffer、dNTP Mixture、IPTG 及DL2000 DNA marker等购自大连宝生物工程公司;琼脂糖、蛋白胨、酵母以及SYBR Green荧光染料等购自基因公司;PCR产物回收纯化试剂盒购自QIAGEN公司;PVDF膜购于Millipore公司;预染蛋白质分子量标准购自Bio-Rad公司;脱脂奶粉购自Sigma公司;StrepTrapTMHP 1 ml柱子购自 GE公司。
1.2 试验方法
1.2.1 密码子优化及引物设计 根据其在Gen-Bank中的ID号AJ427413.2获得其核苷酸和氨基酸序列,确定开放阅读框为156个氨基酸,468碱基对,引入NdeⅠ、PstⅠ、XhoⅠ酶切位点及 Strep TagⅡ。根据原核表达载体大肠杆菌对密码子的偏爱性,同时考虑RNA的二级结构,提高翻译起始效率,利用DNAStar软件进行密码子优化。密码子优化完成后GC含量由44.3%变为47.6%。优化后的全基因序列由上海博尚生物有限公司合成。
1.2.2 Pla a1基因表达载体的构建 Pla a1基因优化后的全编码区序列设计一对引物。引物由上海博尚生物有限公司合成。上游引物为5'-CT-,下游引物 5'-GCCTGCAGAGCACCAAGCAGTTT-3',上下游引物退火温度分别为:68.2℃和69.1℃,黑体下划线部分分别为NdeⅠ、PstⅠ酶切位点。PCR扩增程序为95℃预变性5分钟;95℃变性30秒,63.3℃退火30秒(1℃/touchdown/cycle),72℃延伸30秒,7个循环;95℃变性30秒,57.3℃退火30秒,72℃延伸30秒25个循环;72℃延伸5分钟。对基因公司合成后甘油菌TOP10-Puc57-Pla a1行单克隆菌落PCR,用1%的琼脂糖凝胶电泳检测菌落PCR结果。对鉴定阳性单克隆菌落摇菌培养,提取质粒,NdeⅠ和PstⅠ双酶切,同时对改造过的pET44a质粒载体(载体上含有Strep TagⅡ)NdeⅠ和PstⅠ双酶切位点,获得相同粘性末端,2%的琼脂糖凝胶电泳分别进行目的片段Pla a1及载体pET-44a回收;并进行连接反应,目的片段与载体之比为3,T4 DNA连接酶于4℃连接16小时,构建重组质粒pET44a-Pla a1;将连接产物经KCM(0.5 mol/L KCl,0.15 mol/L CaCl2,0.25 mol/L MgCl2)法转化大肠杆菌感受态JM109;通过氨苄青霉素Amp抗性LB平板筛选,菌落PCR鉴定阳性克隆,并将阳性单克隆过夜培养菌液送上海博尚生物有限公司测序;选取测序正确的质粒,转化高效表达菌株感受态Rosetta,氨苄青霉素Amp抗性LB平板筛选,菌落PCR鉴定阳性克隆,并将阳性单克隆过夜培养菌液送测序,选取测序正确的单克隆菌落诱导蛋白表达。
1.2.3 Pla a1蛋白表达及鉴定 挑取测序阳性重组质粒pET44a-Pla a1的E.coli Rosetta分别接种至含有Amp(50 mg/L)的LB液体培养基(10 g/L胰蛋白胨、5 g/L酵母提取物、10 g/L NaCl)中,37℃振荡培养12小时后按1%的接种量接种到新鲜的含Amp(50 mg/L)、含葡萄糖(4%)的LB液体培养基中培养至OD600在0.6~0.7之间。然后加入终浓度为0.5 mmol/L IPTG诱导剂,于37℃诱导表达4小时。收集菌体,并进行Pla a1表达蛋白的SDSPAGE。电泳方法参照文献[10]进行。取1.5 ml菌液离心收集菌体,加入100 μl 2×上样缓冲液(20 g/L SDS、500 ml/L 甘油、62.5 mmol/L Tris-HCl(pH6.8)、20 ml/L β-巯基乙醇、0.1 g/L 溴酚蓝、30 mmol/L NaCl、1 mmol/L EDTA)充分混合,煮沸5分钟后,12 000 r/min离心SDS-PAGE电泳检测。
1.2.4 鉴定蛋白表达形式 安装上述方法诱导蛋白表达;4℃、10 000 r/min、6分钟收集菌液,按菌体湿重为5ml/1g加结合缓冲液(100 mmol/L Tris-HCl,150 mmol/L NaCl,1 mmol/L EDTA,pH8)充分重悬菌液。冰水浴中超声裂解,裂解5秒停5秒,强度50%,100 W,裂解10分钟;4℃,12 000 r/min,30分钟离心分别收集上清1和沉淀;对上述沉淀再次用相同体积的结合缓冲液重悬洗剂,4℃,12 000 r/min,30分钟离心再次收集沉淀;对沉淀加入相同体积含6 mmol/L尿素结合缓冲液(100mmol/L Tris-HCl, 150mmol/L NaCl, 1 mmol/L EDTA,pH8.0),重悬沉淀,4℃摇床裂解过夜;4℃,18 000 r/min,30分钟离心收集上清2;SDS-PAGE鉴定表达蛋白是否为包涵体形式。
1.2.5 Pla a1表达蛋白的纯化 将变性后蛋白分别在含4 mol/L尿素、2 mol/L尿素、1 mol/L尿素的结合缓冲液各透析12小时。透析后的液体经4℃,18 000 r/min,10分钟收集上清3,准备过柱纯化诱导表达的蛋白。购买于GE公司的Strep Trap HP 1 ml预装柱与AKTA FPLC连接并首先用结合缓冲液按1 ml/min流速平衡柱子,至紫外基线平稳;将上清3上样过柱,并用结合缓冲液洗柱。并收集穿透峰,用洗脱缓冲液洗柱(2.5 mmol/L脱硫生物素100 mmol/L Tris-HCl,150 mmol/L NaCl,1 mmol/L EDTA,pH8.0)并收集洗脱峰。SDS-PAGE蛋白电泳鉴定洗脱峰及穿透峰。
1.2.6 纯化蛋白Western blot鉴定分析 纯化的融合蛋白经SDS-PAGE电泳后,将凝胶上的蛋白质电转移至PVDF膜上,用含50 g/L脱脂奶粉的PBST缓冲液 (8 g/L NaCl 0.28 g/L KCl 0.28 g/L KH2PO4,2.98 g/L Na2HPO4·12H2O,pH7.4;1 ml/L Tween 20)室温封闭2小时后,加入法桐花粉过敏患者血清(一抗1∶1稀释),4℃三维摇床过夜孵育;用PBST在室温下三围摇床上洗4次,每次5分钟;再加入辣根过氧化物酶标记的羊抗人IgE抗体(1∶500),室温孵育1小时,用PBST在室温下三围摇床上洗2次,每次5分钟;再用PBS洗2次,5分钟;用二氨基联苯胺(DAB)底物显色液显色10分钟,检测Pla a1的抗原性。
1.2.7 纯化后的表达蛋白送测序 SDS-PAGE蛋白电泳后,剪取合适大小的PVDF膜,在横流45 mA室温条件下进行电印迹转移,转移时间为28分钟。转印结束,考马斯亮蓝染色(0.1%考马斯亮蓝R-250溶于40%甲醇/1%乙酸)染色30秒,用脱色液(1%醋酸+50%甲醇脱色),脱色后用去离子水充分洗涤,然后剪下待测序的条带,封存在1.5 ml的离心管中送中国科学院上海生命科学研究院生化与细胞研究所蛋白质组研究分析中心测序。
2 结果
2.1 重组表达载体的鉴定 重组载体构建成功后以T7启动子开始,在NdeⅠ和PstⅠ酶切位点之间为目的片段,PstⅠ酶切位点之后为8个氨基酸的Strep TagⅡ、双终止密码子TAATAA及XhoⅠ酶切位点。PET44a-Pla a1重组子转入JM109,对规整化后的菌落进行菌落PCR,结果如图1。1~9为目的片段,菌落PCR目的片段在500 bp左右,符合目标片段大小。质粒测序的结果与原始序列同源性为100%,可以进行下一步原核表达菌株构建。
2.2 Pla a1蛋白表达及鉴定 图2所示,Pla a1在表达菌株Rosetta中的诱导表达,2~4在IPTG浓度为0.5 mmol/L诱导下的表达情况,1为未经IPTG诱导的对照样品。在对照和诱导间(18 kD处)出现明显差别,表明外源基因进行了Pla a1蛋白表达。采用的marker为03-04蛋白marker。
2.3 鉴定蛋白表达形式 在进行蛋白诱导后的收菌,加非变性裂解液,进行超声裂解,裂解后分别取上清和沉淀进行SDS-PAGE电泳。如图3:1为对照,2为诱导,3为超声裂解后上清,4为超声裂解后沉淀。由此可见目的蛋白主要包涵体表达。
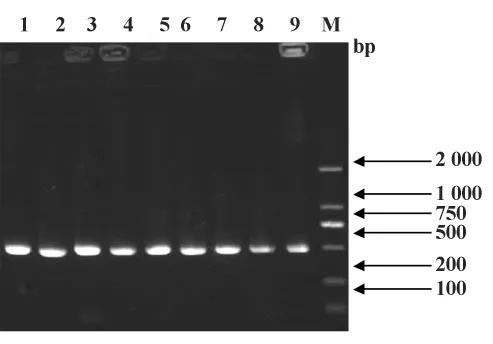
图1 JM109菌落PCR产物电泳结果Fig.1 PCR amplification of the Pla a1 gene
2.4 Pla a1融合蛋白的纯化 利用AKTA FPLC将经过StrepTrap HP 1 ml预装柱所收集的液体分别取100 μl加入20 μl 6 × loading buffer(内含 β-巯基乙醇)煮沸6分钟后,离心(10 000 r/min,5分钟)后取其上清10 μl跑SDS-PAGE胶,如图4:1为穿透峰;2为目的蛋白洗脱峰。1泳带为上样后穿透峰所收集的液体,2泳带为蛋白洗脱时紫外吸收峰大于50 mA时所收集的液体。
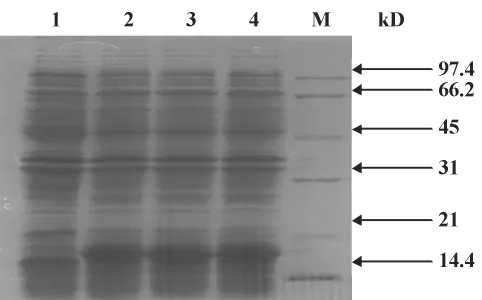
图2 Pla a1基因诱导表达的鉴定Fig.2 Protein expression of Pla a1 gene
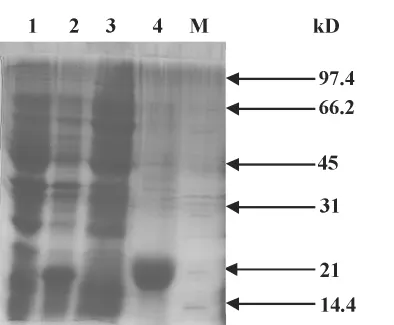
图3 重组蛋白表达形式鉴定Fig.3 Identification of inclusion body protein
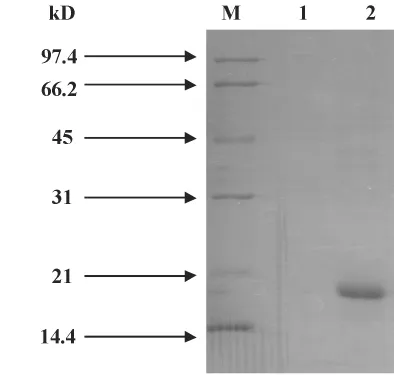
图4 重组蛋白纯化后SDS-PAGE电泳鉴定Fig.4 Identification of the purified recombinant protein by SDS-PAGE
2.5 纯化蛋白Western blot鉴定分析 将诱导纯化后蛋白利用法桐花粉过敏患者血清进行免疫印迹鉴定如图5,泳道1为正常血清对照;2~6所用患者血清均经过ImmunoCAP检测,并且血清中法桐花粉变应原特异性IgE抗体均在二级以上。从图5中可以看到在第2~6泳道内目标大小处有一条清楚的显色条带,表明此目的蛋白具有一定抗原性。
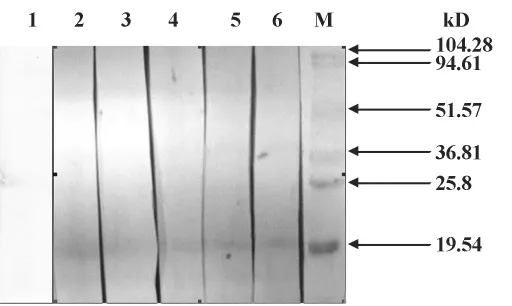
图5 重组Pla a1蛋白的Western blot鉴定Fig.5 Identification of the Pla a1 protein by Western blot
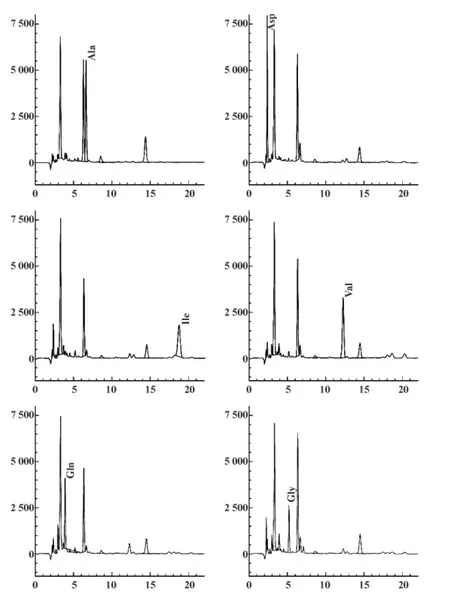
图6 重组蛋白测序结果Fig.6 Result of N-terminal amino acid sequence of protein Pla a1
2.6 纯化后的融合蛋白送测序 纯化后的融合蛋白转PVDF膜后送中国科学院上海生命科学研究院生化与细胞研究所蛋白质组研究分析中心测序。蛋白质N-端1~6个氨基酸序列测序图谱如图6。
3 讨论
大量的花粉流行病学调查资料显示我国以及欧美国家法桐花粉飘逸广泛[1,2,4,11],更多的文献指出法桐花粉已经引起相关花粉症大量出现,如过敏性哮喘、过敏性鼻炎、荨麻疹等[7,9,12-15]。但是对法桐花粉主要变应原鲜有研究,本文对法桐花粉主要变应原进行了表达、纯化及免疫印记鉴定。
大量研究表明,影响外源基因表达效率的重要因素为密码子偏爱性、mRNA稳定性、翻译起始效率和载体选择等[16]。密码子的偏爱性研究在近年来成为热点,如果目的基因的密码子与表达宿主不匹配,则会降低mRNA的翻译效率和稳定性,甚至会造成mRNA翻译的提前终止[17]。本课题通过密码子优化使GC含量由44.3%变为47.6%,在翻译起始区尽量选用最优密码子,并且充分考虑RNA的二级结构,使起始翻译区处于线性开放结构,这样使蛋白质翻译更容易进行。
本课题收集法桐花粉变应原皮试过敏患者血清18例,利用ImmunoCAP检测,9名患者血清法桐花粉变应原特异性IgE抗体检测均在二级以上,利用纯化后的外源目的蛋白与这9名法桐花粉过敏血清进行免疫印记实验,结果显示有5例发生阳性反应,阳性反应率超过50%,经免疫印迹鉴定为法桐花粉主要变应原蛋白,并且证明该外源重组蛋白具有相应的抗原性。
随着分子生物学技术的发展,利用基因工程制备重组花粉主要变应原用于变态反应性疾病的临床诊疗和研究已成为变态反应学科的发展方向之一。重组变应原具有纯度高、产量高、易于标准化、无外源性毒性物质和病原微生物污染的优势。体外试验及皮肤试验的结果均表明,大部分基因重组变应原的变应原活性与天然变应原相当[18]。构建含有变应原编码基因的重组质粒,表达出有功能活性的重组变应原是临床诊断和治疗过敏性疾病的需要,也将为深入研究变应原分子结构和致病机制提供材料。本课题工作为后续低致敏过敏原的改造及变应原核酸疫苗的获得奠定基础。
致谢:本实验在广州医学院第二附属医院,呼吸疾病国家重点实验室,变态反应研究室,过敏反应与临床免疫重点研究室完成,感谢实验室老师、同学对我的帮助、支持。
1 Kosisky S E,Marks M S,Nelson M R.Pollen Aeroallergens in the Washington,DC,Metropolitan Area:A 10-Year Volumetric Survey(1998-2007)[J].Ann Allergy Asthma Immunol,2010;104(3):223-235.
2 Aira M J,Rodriguez-Rajo F J,Fernandez-Gonzalez M et al.Airborne Pollen of Ornamental Tree Species in the NW of Spain[J].Environ Monit Assess,2010;173(1-4):765-775.
3 Ture C,Bocuk H.Analysis of airborne pollen grains in bilecik,Turkey[J].Environ Monit Assess,2009;151(1-4):27-35.
4 Celenk S,Canitez Y,Bicakci A et al.An Aerobiological study on pollen grains in the atmosphere of North-West turkey[J].Environ Monit Assess,2009;158(1-4):365-380.
5 刘光辉,祝戎飞,张 威et al.湖北省气传花粉调查[J].中华临床免疫和变态反应杂志,2007;1(001):22-26.
6 卢家美,孙秀珍,刘 昀et al.西安市气传花粉调查[J].西安交通大学学报(医学版),2010;31(04):472-474,480.
7 Lauer I,Miguel-Moncin M S,Abel T et al.Identification of a Plane Pollen Lipid Transfer Protein(Pla a 3)and its immunological relation to the Peach Lipid-Transfer Protein,Pru P 3[J].Clin Exp Allergy,2007;37(2):261-269.
8 Asturias J A,Ibarrola I,Eraso E et al.The major platanus acerifolia pollen allergen Pla a 1 has sequence homology to invertase inhibitors[J].Clin Exp Allergy,2003;33(7):978-985.
9 Ibarrola I,Arilla M C,Martinez A et al.Identification of a Polygalacturonase as a Major Allergen(Pla a 2)from Platanus Acerifolia Pollen[J].J Allergy Clin Immunol,2004;113(6):1185-1191.
10 Laemmli U K.Cleavage of structural proteins during the assembly of the head of bacteriophage T4[J].Nature,1970;227(5259):680-685.
11 Celenk S,Bicakci A,Tamay Z et al.Airborne Pollen in European and Asian parts of istanbul[J].Environ Monit Assess,2009;164(1-4):391-402.
12 Subiza J,Cabrera M,Valdivieso R et al.Seasonal asthma caused by Airborne Platanus Pollen[J].Clin Exp Allergy,1994;24(12):1123-1129.
13 Pazouki N,Sankian M,Leung P T et al.Identification of cyclophilin as a novel allergen from platanus orientalis pollens by mass spectrometry[J].J Biosci Bioeng,2009;107(2):215-217.
14 Fernández-González D,González-Parrado Z,Vega-Maray A M et al.Platanus Pollen Allergen,Pla a 1:Quantification in the atmosphere and influence on a sensitizing population[J].Clinical& Experimental Allergy,2010;40(11):1701-1708.
15 Varela S,Subiza J,Subiza J L et al.Platanus pollen as an important cause of pollinosis[J].J Allergy Clin Immunol,1997;100(6 Pt 1):748-754.
16 Baneyx F.Recombinant protein expression in Escherichia Coli[J].Current Opinion in Biotechnology,1999;10(5):411-421.
17 郑彬琼.大肠杆菌同义密码子偏好性概述[J].硅谷,2009;(01):3,24.
18 Nikolaizik W H,Weichel M,Blaser K et al.Intracutaneous tests with recombinant allergens in cystic fibrosis patients with allergic bronchopulmonary aspergillosis and aspergillus allergy[J].Am J Respir Crit Care Med,2002;165(7):916-921.
