不同碳源对干酪乳杆菌LC2W胞外多糖组成影响的研究
2012-09-11刘翠平吴正钧
刘翠平,吴正钧
光明乳业股份有限公司乳业生物技术国家重点实验室,上海 200436
乳酸菌产胞外多糖(exopolysaccharide,EPS)指乳酸菌(lactic acid bacteria,LAB)在生长代谢过程中分泌到细胞壁外的黏多糖或荚膜多糖。LAB EPS是由许多重复单位构成的带有分支结构的多糖,其独特的物理流变学特性、使用安全性以及生理活性,使它在食品和非食品工业倍受青睐,尤其是它在医药领域所具有的巨大应用潜能正日益引起人们的广泛关注。如:可以用做增稠剂、凝胶剂和稳定剂,能降低胆固醇,抗癌,抗溃疡,改善肠道微生态环境,增强免疫力或作为益生元等[1-5]。
LAB EPS的活性与结构密切相关,乳酸菌胞外多糖的结构分析对预测其生物活性功能,进一步研究结构与功能的关系非常重要。培养基组成尤其是碳源的改变对胞外多糖分子量、单糖组成和生物学活性均可能有影响[6,7],目前的研究还存在争议。本文利用HPLC、气相色谱、紫外光谱和红外光谱等测试手段对两种碳源获得的EPS均一多糖进行初步结构分析,为干酪乳杆菌LC2W EPS的进一步研究和利用以及分析构效关系奠定理论基础。
1 材料与方法
1.1 材料与仪器
均一多糖G1、G2(以葡萄糖为碳源经LC2W发酵分离纯化制备)[8-10];均一多糖 L1、L2(以乳糖为碳源经LC2W发酵分离纯化制备);

表1 实验主要仪器Table 1 Experimental apparatus and equipments list
1.2 实验方法
1.2.1 G1、G2 与L1、L2 的纯度鉴定及相对分子量比较
高效液相色谱(HPLC):将不同分子量的标准Dextran相继进样,记录保留时间TR,以TR为横坐标,LgM为纵坐标绘制标准曲线,得出分子量与保留时间TR的回归方程。待测样品按上述步骤进样,根据所得TR,通过回归方程计算样品的相对分子量。高效液相色谱条件:色谱仪Waters 2690;检测器Waters 2410示差折光检测器;色谱柱 Waters UltrahydrogelTM Linear(φ7.8mm ×300 mm ID)两柱串连;流动相0.1 mol/L NaNO3;柱温45 ℃;流速0.9 mL/min;进样量 20 μL
1.2.2 紫外光谱(UV)分析比较
将多糖配成0.5 mg/mL溶液,在波长190~400 nm范围内进行紫外光谱扫描,观察在波长280 nm和260 nm处有无吸收峰,确定多糖样品中是否含有蛋白质或核酸。
1.2.3 红外光谱分析比较
将干燥多糖样品2.0 mg与KBr研磨压片,在4000~500 cm-1区域内进行红外光谱扫描。
1.2.4 单糖组成及其摩尔比比较
取待分析多糖样品,每15~20 mg加入2 mL 1mol/L H2SO4封管105℃水解3 h,BaCO3中和,过滤去掉沉淀,真空冻干,而后进行衍生化。
衍生物的制备(糖腈乙酸酯衍生法):称取10 mg固体水解糖样、10 mg盐酸羟胺和1~2 mg内标,加入0.5 mL吡啶,90℃放入水中加热反应30 min并振荡。取出后冷至室温,加入0.5 mL醋酸酐,在90℃下继续反应30 min进行乙酰化,反应产物直接用于气相色谱分析。
气相色谱检测条件如下:仪器 日本岛津GC-14A;色谱柱 DB-1701 30M I.D.0.53 mm ×1.5 μm;检测器FID(氢火焰离子化检测器);汽化室温度260℃;检测器温度260℃;程序升温120(2)195(0.1)/10-240(10)/3;载气压力 (N2):0.80 kg/cm2;燃气压力(H2):0.65 kg/cm2;助燃气压力(空气):0.50 kg/cm2;分流比 30∶1;进样量 0.8 μL。
2 结果与讨论
2.1 G1、G2 与 L1、L2 的相对分子量比较
干酪乳杆菌LC2W分别以两种碳源发酵产粗多糖,经离子柱和凝胶柱分离纯化后得到的中性糖G1、G2和L1、L2经过HPLC后峰形均呈单一对称峰,都是均一多糖。通过HPLC测得的各均一组分平均相对分子量如表2所示。

表2 不同碳源来源多糖平均相对分子量比较Table 2 Compare of average molecular weight
不同碳源获得的多糖组分分子量存在明显差异,以乳糖为碳源得到的中性糖两组分相对分子量均高于以葡萄糖为碳源获得的中性糖两组分的相对分子量,与Sepharose CL-6B凝胶柱层析层析出峰时间差异现象相吻合。可能改变碳源影响了其单糖组成。
2.2 紫外光谱分析比较
G1、L1和G2、L2的全波长扫描结果见图1、图2,光谱显示,四组分仅在190 nm处有一吸收峰,这是多糖特征性的紫外吸收峰,在260 nm和280 nm处均无明显吸收,说明均不含有核酸、蛋白质[11]。
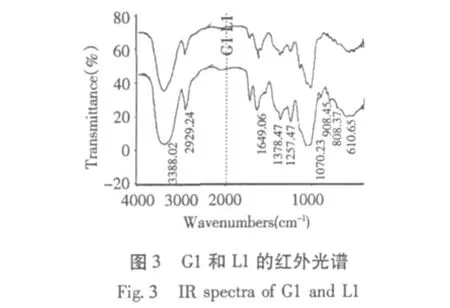
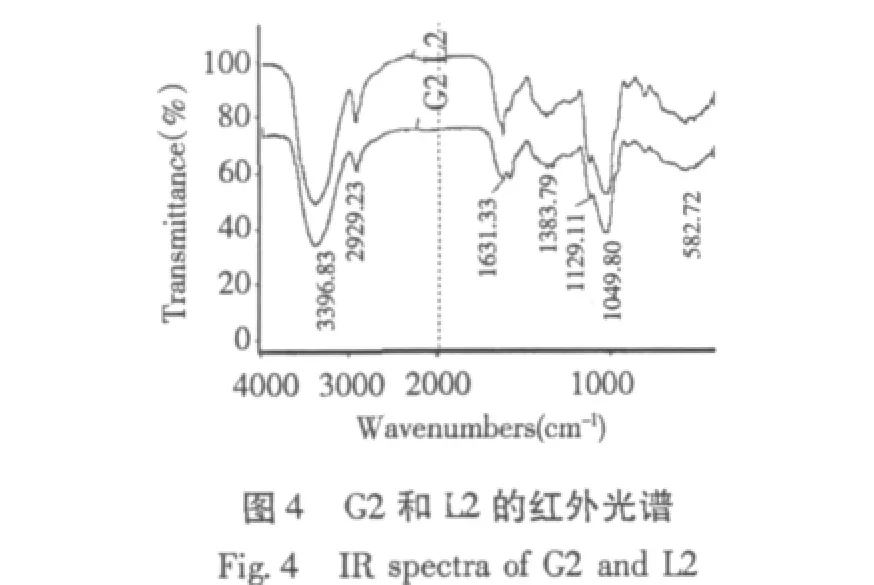
2.3 红外光谱分析比较
G1、L1 和 G2、L2 的红外光谱见图3、图 4,红外光谱显示,不同碳源发酵分离纯化得到的多糖四组分在4000 cm-1~500 cm-1范围内都具有糖类的特征吸收峰。3700 cm-1~3000 cm-1出现的宽吸收峰是O-H的伸缩振动,表明各多糖均存在分子内或分子间的氢键。2930 cm-1和1385 cm-1的吸收峰由C-H伸缩振动和弯曲振动引起,1400 cm-1~1200 cm-1所看到不太尖的吸收峰是C-H的变角振动,1200 cm-1~1000 cm-1区域吸收峰为各种多糖的特征吸收峰,主要两种C-O伸缩振动引起,其中之一是C-O-H的,另一种是糖环 C-O-C的[12]。
初步可以看出不同来源多糖组分结构中出现了强度不同的特征吸收峰,在谱峰形状及特征吸收波段上有些差别,表明其在结构组成上可能有差异。
2.4 单糖组成及其摩尔比比较
分析多糖样品单糖组成常用气相色谱法,气相色谱法有选择性强、分辨力好、灵敏度高以及分析速度快等优点[12]。本试验采用气相色谱法对两种不同碳源获得的多糖组分进行单糖组成分析,标准样品经糖腈乙酸酯衍生化,在1.2.4检测条件下测得的各标样气相色谱图见图5。

图5 标准单糖的气相色谱图Fig.5 Gas chromatogram of standard sample
样品经酸水解和衍生化处理后,经气相色谱检测,结果如表3所示。根据保留时间确定样品单糖组成成分,单糖含量(%)=Ai/AIi*AIS/As*WS/WIS*WIi/W*100
Ai表示某单糖样品峰面积;
AIi表示样品中加入的肌醇内标峰面积;
AIS表示标样中加入的肌醇内标峰面积;
As 表示某单糖标样峰面积;
WS表示某单糖标样的质量mg;
WIS表示标样中加入的肌醇内标的质量mg;
WIi表示样品中加入的肌醇内标的质量mg;
W 表示称取多糖样品的质量mg。
多糖的单糖摩尔比以计算得到的单糖质量除以其摩尔质量再计算得到,由图5及表3可知,G1和L1单糖组成成分相同,而且相对简单,都是主要由鼠李糖、半乳糖和葡萄糖组成,另外含有少量的甘露糖,但对应单糖组分百分含量不同,G2和L2单糖组成比G1和L1复杂,主要由甘露糖、葡萄糖和半乳糖组成,另外含有很少量的鼠李糖、阿拉伯糖和木糖,对应单糖组分百分含量不同。
四组分的主要单糖组成摩尔比如下:
G1——鼠李糖∶葡萄糖∶半乳糖 =3.07∶3.29∶1;
L1——鼠李糖∶葡萄糖∶半乳糖 =2.84∶6.4∶1;
G2——甘露糖∶葡萄糖∶半乳糖 =17.24∶6.03∶1;
L2——甘露糖∶葡萄糖∶半乳糖 =28.71∶5.98∶1。
分析表明,不同碳源获得的多糖中性糖两组分主要单糖组成摩尔比上有一定差异。G1和L1单糖组成相同但摩尔比不同,G2和L2也出现同样现象,结合高效液相测定的平均相对分子量分析,改变碳源改变了干酪乳杆菌LC2W合成胞外多糖的单糖组成摩尔比,可能也是造成其分子量差异的原因。

表3 G1、L1、G2、L2的单糖组成及质量分数(%)Table 3 Sugar composition and mass fractions of G1,L1,G2,L2(w/w%)
3 结论
将碳源以乳糖替换,其余条件都不变,分离纯化中性多糖相应组分,并从分离纯化结果,分子量,单糖组成及摩尔比,紫外和红外光谱分析几方面进行了分析比较,结果表明,改变碳源,EPS的分子结构、分子量和单糖组成都发生了变化。改变碳源对干酪乳杆菌LC2W胞外多糖功能活性影响及具体构效关系还需进一步探讨。
1 Guo BH(郭本恒).Probiotics(益生菌).Beijing:Chemical Industry Press,2004.
2 Mou GQ(牟光庆),Li X(李霞),Li YX(李瑶喜),et al.Enhancement Effect of Extracellular Polysaccharides Produced by Lactic Acid Bacteria on Immunologic Function of Mice.Food Sci(食品科学),2009,30:260-262.
3 Dong W(董伟),Tang YJ(唐彦君),Liu N(刘宁),et al.Effects of exopolysaccharide from Lactobacillus plantarum on secretion of dendritic cells in the mouse marrow.Chin Dairy Ind(中国乳品工业),2011,39(6):14-16.
4 Yang JB(杨洁彬),Guo XH(郭兴华),et al.Lactobacillus-Biological Basis and Application(乳酸菌-生物学基础及应用).Beijing:China Light Industry Press,1996.l-3.
5 Sutherland IW.Bacterial exopolysaccharides.Adv Microbiol Physiol,1972,8:143-212.
6 Van Den Berg DJC,Robijn GW,Janssen AC.Production of a novel extracellular polysaccharide by Lactobacillus sake 0-1 and characterization of the polysaccharide.Appl Environ Microbiol.1995,61:2840-2844.
7 Gassem MA,Schmidtand KA,Frank JF.Exopolysaccharide production by Lactobacillus delbrueckii ssp.Bulgaricus.J Food Sci,1997,62:171-173,207.
8 Liu CP(刘翠平),Guo BH(郭本恒),Wu ZJ(吴正钧),et al.Effects of nutrition factors on the exopolysaccharide production by Lactobacillus casei LC2W(营养因子对干酪乳杆菌LC2W胞外多糖合成的影响).Ind Microbiol(工业微生物),2008,38(2):11-14.
9 Liu CP(刘翠平),Wu ZJ(吴正钧),Wang YY(王荫榆),et al.Optimization of Culture Conditions for the Production of Exopolysaccharides by Lactobacillus casei LC2W Using Response-surface Methodology(响应面法优化干酪乳杆菌LC2W合成胞外多糖的培养条件).Food Ferment Ind(食品与发酵工业),2008,34(12):85-89.
10 Liu CP(刘翠平),Ye L(叶蕾),Wu ZJ(吴正钧),et al.Optimization of Preparation Procedures for Exopolysaccharides from Lactobacillus casei LC2W(干酪乳杆菌LC2W胞外多糖提取工艺研究).Acta Agric Boreali-Sinica(华北农学报),2008,23(supplement):118-121.
11 Si L(石磊),Fu YL(傅佑丽),Chen KS(陈靠山),Separation,Purification and Component Research of a Polysaccharide from the Roots of Cudrania tricuspidata(Carr.)Bur..J Shandong Univ,Nat Sci(山东大学学报,理学版),2007,42(5):74-78.
12 Zhang WJ(张惟杰).Biochemistry Research Technique of Glycoconjugates(2ndEdition)(糖复合物生化研究技术,第二版),Hangzhou:Zhejiang University Press,2003.
