苏北滨海湿地互花米草两种繁殖体与本地种竞争能力分析*
2012-09-06陈正勇王国祥刘金娥徐伟伟
陈正勇,王国祥,刘金娥*,徐伟伟,王 刚
(1.南京师范大学 地理科学学院,江苏 南京210046;2.江苏省环境演变与生态建设重点实验室,江苏 南京210046;3.安徽师范大学 环境科学学院,安徽 芜湖241003)
苏北滨海湿地互花米草两种繁殖体与本地种竞争能力分析*
陈正勇1,2,王国祥1,2,刘金娥1,2*,徐伟伟1,2,王 刚3
(1.南京师范大学 地理科学学院,江苏 南京210046;2.江苏省环境演变与生态建设重点实验室,江苏 南京210046;3.安徽师范大学 环境科学学院,安徽 芜湖241003)
在碱蓬滩选取互花米草斑块,由外向内分为外围(A)、外侧(B)、内侧(C)、中心(D)四个取样点,研究互花米草实生苗、分蘖苗与本地植物碱蓬、芦苇的相对竞争力,探讨互花米草在碱蓬滩地入侵扩张的趋势。结果表明:1)在斑块B点处,互花米草实生苗与碱蓬相对竞争力高于互花米草分蘖苗与碱蓬相对竞争力;在斑块C点处,互花米草实生苗与碱蓬相对竞争力低于互花米草分蘖苗与碱蓬相对竞争力;多样性指数分析表现为:B点>C点>A点>D点;在互花米草斑块B点处重要值大小表现为:互花米草实生苗>碱蓬>芦苇实生苗>互花米草分蘖苗,在互花米草斑块C点处重要值大小表现为:互花米草分蘖苗>芦苇实生苗>互花米草实生苗>碱蓬。因此,碱蓬滩互花米草斑块主要是通过实生苗入侵新生境,向外扩张,通过互花米草分蘖苗维持种群的稳定。2)在斑块B点,芦苇实生苗与互花米草实生苗的相对竞争力相当,大于互花米草分蘖苗的相对竞争力,但在斑块C点处,互花米草分蘖苗高于芦苇实生苗。碱蓬与本地种芦苇也产生了竞争关系,并很有可能被芦苇和互花米草共同取代。3)互花米草实生苗与碱蓬的相对竞争力与滩涂微生境土壤含水量、土壤全盐量负相关(P<0.01),与土壤pH正相关(P<0.01);互花米草分蘖苗与碱蓬的相对竞争力与土壤含水量、土壤全盐量正相关(P<0.05),与土壤pH负相关(P<0.01)。碱蓬滩斑块处的含水量、盐度适合互花米草的定居、扩张。
外来种;繁殖方式;扩张趋势;种间竞争;生物入侵
互花米草(Spartina alterniflora)由于耐盐耐淹、繁殖力强等特点[1],在世界上多处引种地扩张迅速,成为世界范围的入侵种[2]。互花米草的入侵和爆发机制主要涉及3个方面[3]:物种的入侵力、生态系统的可入侵性以及在恰当的时间提供的入侵通道。已有研究阐明了一些外来种入侵的机制,目前研究者已经普遍用定量的方法,从外来入侵种的生态幅、繁殖传播特性、种群遗传结构等多个视角做了不少工作,研究表明互花米草对环境因子适应幅度较广,比土著种更适应当地自然环境;具有有性和无性两种繁殖方式;种子可远距离传播;遗传多样性低减少了种内竞争;新栖息地选择压力产生新的有利入侵的性状等[3-4]。但尚不能对入侵种的特性作出普适性的结论[5]。
江苏沿海拥有我国最大的滩涂湿地,互花米草自1980年引入苏北滩涂,扩张迅速,对当地生态系统的结构和功能产生较强的干扰,主要表现为改变潮间带地形,阻碍河道畅通[6];竞争取代土著植物[7];改变大型底栖无脊椎动物群落的结构,威胁滩涂养殖业[8-9];降低昆虫、鸟类以及根际微生物的多样性[10-11]。碱蓬是苏北滩涂的典型本地种,根据我们在盐城海滨湿地跟踪观测,互花米草以斑块形式出现,和碱蓬已经形成了交错带,与芦苇、碱蓬伴生共存。未来竞争结果如何?互花米草是否有可能替代本地种?两种繁殖体哪一种在扩张中更具有竞争力?本研究通过分析互花米草实生苗和分蘖苗与盐城滨海湿地土著植物的相对竞争能力,探讨互花米草在江苏盐城滨海湿地的扩张模式和趋势。
1 研究材料与研究方法
1.1 研究区概况
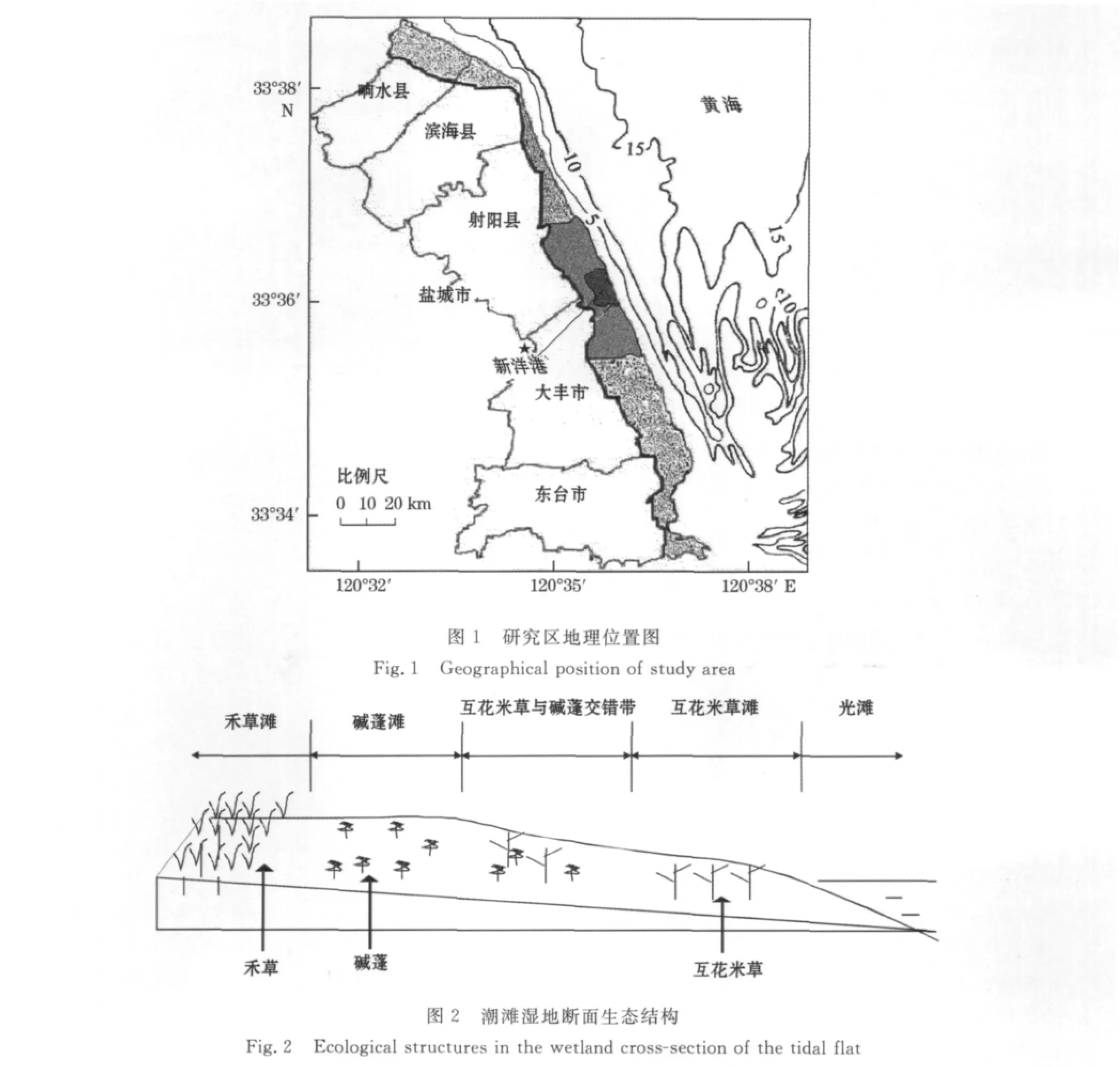
研究区新洋港位于33°32′~33°40′N,120°30′~120°40′E之间(图1),高程范围为:2~2.6m。研究区受人类干扰较少,基本保留海岸带原生植被演替序列。由海向陆依次为光滩、互花米草(S.alterniflora)滩、盐地碱蓬(Suaeda salsa)滩及禾草(藨草Scirpus trqueter、獐毛Aeluropus littoralis、白茅Imperata cylindrica、芦苇Phragmites australis)滩(图2)。
1.2 数据收集及处理
2010-07,于碱蓬滩选取互花米草斑块,斑块由外向内分为外围、外侧、内侧、中心(图3),分别以A,B,C,D表示,4个点的物种组成分别为:碱蓬+芦苇、碱蓬+芦苇+互花米草、碱蓬+芦苇+互花米草、互花米草。每个采样点中分别随机设置3个样方(25cm×25cm),采取植物及土壤。测量每株植物的生长与存活指标(密度、株高、茎长、叶长、叶数、枯叶数)以及地上生物量参数。株高以克隆系中最高个体为准。叶长为第三全展叶片的长度.测量后,将互花米草地上部分,80℃烘干称恒重。
每取样点处用环刀法取3处土壤剖面0~20cm表层样品,混合,带回实验室。土壤化学指标测定方法[12]如下:采用烘干法测定土壤含水量,样品待自然风干后研磨,过150μm孔径筛;采用电导法测定土壤全盐;采用电极电位法测定土壤pH值。
1.3 竞争力研究
1.3.1 相对竞争力系数
采用de Wit的牧草竞争模型[12]来计算牧草种间竞争的强度,分析互花米草与碱蓬的种间竞争关系,即:


图3 斑块取样点位置示意图Fig.3 A schematic map of the patch-sampling sites
式中,Rij为互花米草对碱蓬的相对竞争系数(Relative Competitive Coefficient),Yij为互花米草与碱蓬混植时互花米草种的单株产量,Yji为互花米草与碱蓬混植时碱蓬的单株产量,Yii为互花米草单植时互花米草的单株产量,Yjj为碱蓬单植时碱蓬的单株产量。
1.3.2 生物多样性指数
1)Shannon-Wiener指数[13]:是种类数和种类中个体分配上的均匀度的综合指标。其公式为

式中,ni为第i个种的个体数目;N为群落中所有种的个体总数。
2)Pielou均匀度指数:反映种间个体分布的均匀性。其公式为

式中,Hmax=lnS为最大的种类多样性;S为总的种类数。
3)Margalef物种丰富度指数:

1.3.3 重要值
重要值(IV)计算:

式中,RDE为相对密度,是群落中某种植物的密度占群落总密度的百分比,它在一定程度上决定着种群的能流、生产力及资源利用;RCO为相对盖度,是群落中某种植物的盖度占总盖度的百分比,它可以反映植物种群在地面上的生存空间,在一定程度上是植物利用环境及影响环境程度的反映;RFE为相对频度,是群落中某种植物出现的频度占群落植物出现总频度的百分比。重要值是评价某一植物种群在群落中作用的综合性数量指标。
1.4 统计分析
采用SPSS 11.5软件进行数据处理,选择单因素方差分析(One-wayANOVA)进行差异显著性检验。
2 结果与分析
2.1 生境特征
互花米草斑块的4个取样点之间的土壤全盐和土壤含水量均呈显著性差异(n=6,P<0.05),土壤pH无显著性差异。各取样点中,土壤含水量表现为:D点>C点>B点>A点。有碱蓬生长的取样点处的土壤全盐量低于其它取样点土壤全盐。互花米草可能通过泌盐作用将土壤中盐分泌出,保留在叶片之上,从而降低了土壤盐度,互花米草比碱蓬能够有效的改变土壤环境的盐浓度,在碱蓬滩互花米草斑块中,有碱蓬生长的取样点处的土壤全盐高于无碱蓬有互花米草生长的取样点出的土壤全盐(表1)。

表1 碱蓬滩互花米草斑块各取样点之间的生境指标变化(平均值±标准误差)Table 1 Changes in habitat index among different sampling sites in the S.alterniflora patch in the Suaedaflat(mean±standard error)
2.2 植株生长特征
2.2.1 密度
随着取样点位置斑块由外向内推移,碱蓬、芦苇的密度显著减少(图4),A点碱蓬密度是C点碱蓬密度的12.71倍,A点芦苇是C点芦苇密度的1.97倍。而互花米草实生苗主要分布在斑块的边缘,由斑块外向内推移,逐渐减少,互花米草分蘖苗的密度是由斑块外向内推移逐渐减少,斑块D点处互花米草密度是B点密度的8.57倍。

图4 各取样点不同植物的密度Fig.4 Changes in habitat index among different sampling sites
2.2.2 生长指标的变化趋势
调查结果发现(图5),3个斑块植物生长指标均表现为一致的变化趋势:碱蓬滩互花米草斑块中,本地种碱蓬、芦苇互花米草(实生苗、分蘖苗)生长趋势(株高、茎长、叶长、叶数、枯叶数)表现为从外到内先下降再升高,整体表现为升高的趋势。

图5 不同取样点碱蓬、芦苇和互花米草生长变化Fig.5 Growth indices of Suaeda,reed and S.alternifloraat different sampling sites
2.2.3 生物量
实验结果表明(图6),本地种碱蓬和芦苇生物量表现为由取样点从外到内逐渐降低,互花米草生物量逐渐升高。碱蓬滩互花米草斑块中各取样点之间植被生物量均成显著性差异(n=3,P<0.05)。

图6 各取样点不同植物的生物量Fig.6 Biomass of different plants at the different sampling sites
2.3 互花米草分蘖苗、实生苗与土著种间竞争特征
2.3.1 竞争指数研究
由于在斑块的A点和D点取样点处物种单一(表1),分别为碱蓬+芦苇和互花米草,故仅计算B点与C点处互花米草实生苗、互花米草分蘖苗、碱蓬、芦苇之间的相对竞争力。在种间,根据RCC的评价标准,R>1时,i种竞争大于j种竞争;R<1时,i种竞争小于j种竞争。结果表明,在斑块B点中(表2),互花米草实生苗的相对竞争力略大于碱蓬,互花米草分蘖苗的相对竞争力则小于碱蓬,互花米草实生苗的相对竞争力与芦苇实生苗相当,而互花米草分蘖苗的相对竞争力则小于芦苇实生苗,芦苇实生苗的相对竞争力则大于碱蓬;在斑块C点中(表3),互花米草实生苗的相对竞争力与碱蓬相当,互花米草分蘖苗的相对竞争大于碱蓬,互花米草实生苗的相对竞争力小于芦苇实生苗,而互花米草分蘖苗的相对竞争力大于芦苇实生苗,实生苗芦苇的相对竞争力则大于碱蓬。
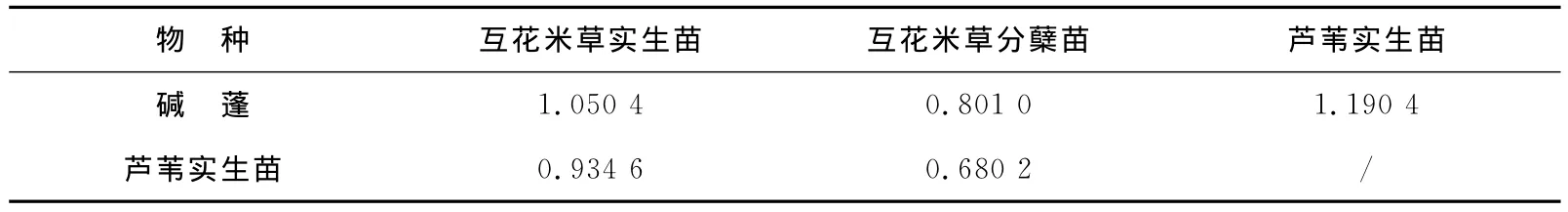
表2 斑块B点碱蓬、芦苇和互花米草相对竞争力Table 2 Relatively competitive abilities of Suaeda,reed and S.alternifloraat site B

表3 斑块C点碱蓬、芦苇和互花米草相对竞争力Table 3 Relatively competitive abilities of Suaeda,reed and S.alternifloraat site C
2.3.2 生物多样性指数
从香农指数和均匀度指数看出(表4),样地的互花米草斑块中,A点和D点的物种单一,在B点和C点中的物种多样性相对丰富,较为均匀。互花米草斑块中,B点的Shannon-Wiener指数值是最高,B点处于斑块边界处,与碱蓬形成了竞争关系,此处的互花米草植株株高相对较小,为碱蓬、芦苇等植物提供了较多的生态位。同时可以发现,碱蓬滩互花米草斑块在B点和C点的丰富度较其他两个取样点丰富(表4),这可以说明,互花米草在碱蓬滩处与碱蓬、芦苇形成了激烈的竞争关系。

表4 斑块Shannon-Wiener指数Table 4 Shannon-Wiener indices at different sampling sites
2.3.3 互花米草斑块植物重要值
从结果发现,在斑块B点处(表5),以互花米草实生苗的重要值最大,斑块B点处主要存在互花米草、碱蓬、芦苇三种植物,互花米草实生苗的重要值在160以上,占绝对优势种的地位,而互花米草分蘖苗的重要值则是最低的;在斑块C点处(表6),以互花米草分蘖苗的重要值最大,互花米草分蘖苗的重要值在190以上,占绝对优势种的地位,而碱蓬的重要值则是最低的。但在斑块A点处,无互花米草的存在,碱蓬保持着优势种地位。这可以说明,在碱蓬、互花米草交错区,互花米草在竞争中处于优势地位,可以成功取代碱蓬成为优势种,这与互花米草能够耐碱、耐旱有着一定的关系。

表5 斑块B点处植物重要值Table 5 Significant value of plants at site B

表6 斑块C点处植物重要值Table 6 Significant value of plants at site C
3 讨 论
3.1 生态系统的可入侵性
互花米草对环境的适应性和耐受能力很强,具有广阔的地理气候范围[14]。相关研究表明[15],土壤水分条件适中有利于互花米草的生长扩张以占领有利的资源环境,而土壤水分条件低则抑制互花米草的生长繁殖,影响其种群延续。互花米草对盐胁迫的高抗性与其多途径的适应机制有关。在盐生环境中互花米草主要采取拒盐的策略,其次才是泌盐和聚盐[16]。相对竞争力与生境特征相关性分析结果表明(表7),互花米草实生苗与碱蓬相对竞争力与土壤含水量、土壤全盐在α=0.01水平上呈负相关,与土壤pH在α=0.01水平上呈正相关;互花米草分蘖苗与碱蓬相对竞争力与土壤含水量、土壤全盐在α=0.01水平上呈正相关,与土壤pH在α=0.01水平上呈负相关;互花米草分蘖苗与芦苇实生苗相对竞争力与土壤含水量、土壤pH在α=0.05水平上呈正相关;芦苇实生苗与碱蓬相对竞争力与土壤含水量、土壤全盐在α=0.05水平上呈正相关。互花米草实生苗与芦苇实生苗相对竞争力与生境特征无相关性。

表7 土壤环境因子与植物相对竞争力间的相关系数Table 7 Correlation coefficients of the environmental factors of soil to the competitive ability of plants
综合结论得出,碱蓬滩互花米草斑块由外至内不断充足的水分条件有利与互花米草分蘖苗保障和维持斑块种群的稳定,互花米草实生苗入侵新生境,并且为芦苇实生苗的定居创造了条件,不断变高的盐度一定程度上抑制了碱蓬种群的生长,而互花米草则由于其特有的生长策略在盐生环境竞争中占优势。
野外观察发现,由于海潮的侵蚀作用,形成了一条由海向陆的潮沟。盐沼面上的潮沟网系具有输送水沙物质及能量(潮流、波浪等)的功能[17],我们推断,互花米草种子可能通过潮沟的水动力作用,顺着潮沟水流将种子带到碱蓬滩处,并在适当的生态环境中定居成功,形成现有斑块。
3.2 互花米草的入侵力
我们可以推断,在碱蓬滩处,互花米草斑块主要是通过实生苗入侵新生境,向外扩张,通过互花米草分蘖苗保障和维持斑块种群的稳定,这主要是取决于互花米草极高的繁殖能力。种子繁殖是互花米草进入新生境的一种重要方式[18-19]。互花米草有性繁殖的贡献主要体现在种群扩散和拓殖[20],产生的大量种子为互花米草种群的快速扩张提供了源泉[5]。对C和D处取样点的互花米草分蘖苗生长指标(株高、茎长、叶长、叶数、枯叶数)的调查发现,C和D点的互花米草分蘖苗生长显著高于B点的互花米草分蘖苗和种子苗,并且实生苗在C点和D点的密度非常的低。由此可见:互花米草在有性繁殖产生的大量种子扩散定居的同时,新形成的克隆也通过极强的萌蘖能力和根状茎快速生长来扩大种群。营养繁殖特征可有效保障种群的维持和更新,这也为其种群快速扩散和爆发提供了可能[21]。
斑块B点处,互花米草实生苗和碱蓬的相对竞争力高于互花米草分蘖苗与碱蓬之间的相对竞争力,并且物种重要值计算结果也表明互花米草实生苗的重要值是最大,显著高于碱蓬、互花米草分蘖苗;而在斑块C点,互花米草分蘖苗和碱蓬的相对竞争力高于互花米草实生苗与碱蓬之间的相对竞争力,并且物种重要值计算结果也表明互花米草分蘖苗的重要值是最大的。随着从斑块外向内推移的过程中,互花米草分蘖苗与碱蓬和、芦苇的相对竞争力逐渐增大,而实生苗的相对竞争力则逐渐减小。而在碱蓬滩处芦苇实生苗的相对竞争力也是高于碱蓬的,并在斑块B点处,芦苇实生苗与互花米草实生苗的相对竞争力相当,大于互花米草分蘖苗的相对竞争力,但在斑块C点处,互花米草分蘖苗的相对竞争力大于芦苇实生苗。我们可以推断,碱蓬与本地种芦苇也产生了竞争关系,并很有可能被芦苇和互花米草共同取代。
4 结 论
通过对碱蓬滩处互花米草斑块内互花米草实生苗和分蘖苗的扩张及与土著植物的相互竞争关系研究我们可以得到以下结论:
1)潮沟为互花米草种子向碱蓬滩扩张提供通道,并为其成功定居提供水分条件,提高了碱蓬滩的可入侵性,为互花米草的定居提供了;
2)在碱蓬滩,互花米草主要是通过实生苗入侵新生境,向外扩张,通过互花米草分蘖苗保障和维持斑块种群的稳定;
3)苏北海滨湿地本地种碱蓬,可能被互花米草和芦苇共同取代,形成以互花米草与芦苇为优势种的群落。
(References):
[1] QIN H,YAO Y H,LI H L.Utilization potential of Spartina alterniflora Loisel as a bio-energy source[J].Chinese Journal of Ecology,2008,27(7):1216-1220.清华,姚懿函,李红丽.互花米草生物质能利用潜力[J].生态学杂志,2008,27(7):1216-1220.
[2] DENG Z F,AN S Q,ZHI Y F,et al.Preliminary studies on invasive model and outbreak mechanism of exotic species,Spartina alterniflora Loisel[J].Acta Ecologica Sinica,2006,26(8):2678-2686.邓自发,安树青,智颖飙,等.外来种互花米草入侵模式与爆发机制[J].生态学报,2006,26(8):2678-2686.
[3] VITOUSEK P M,WAKLE L R,WHITEAKER L D,et al.Biological Invasion by Myrcia fayaalters ecosystem development in Hwaaii[J].Science,1987,238:802-804.
[4] KOLAR C S,LODGE D M.Progress in invasion biology:Predicting invaders[J].Trends in Ecology & Evolution,2001,16(4):199-204.
[5] HOLWAY D A.Competitive mechanisms underlying the displacement of native ants by the Invasive argentine ant[J].Ecology,1999,80(1):238-253.
[6] WANG A J,GAO S,JIA J J.Impact of Spartina alterniflora on sedimentary and morphological evolution of tidal salt marshes of Jiangsu,China[J].Acta Oceanologica Sinica,2006,28(1):92-99.王爱军,高抒,贾建军.互花米草对江苏潮滩沉积和地貌演化的影响[J].海洋学报,2006,28(1):92-99.
[7] CALLAWAY J C,JOSSELYN M N.The introduction and spread of smooth cordgrass(Spartina alterniflora)in south San Francisco Bay[J].Estuaries,1992,15:218-226.
[8] CHEN Z Y,FU C C,WANG H Y,et al.Effects of Spartina alternifloraInvasions on the Benthic Macro-Onvertebrates Community at Dongtan of Chongming Salt Marsh,the Yangtze River Estuary[J].Wetland Science,2005,3(1):1-7.陈中义,付萃长,王海毅,等.互花米草入侵东滩盐沼对大型底栖无脊椎动物群落的影响[J].湿地科学,2005,3(1):1-7.
[9] XU X J,WANG H,YOU W H,et al.A study on fluctuation of zoobenthic community in chongming dongian spartina alterniflora Liusel[J].Transactions of Oceanology and Limnology,2006,(2):89-95.徐晓军,王华,由文辉,等.崇明东滩互花米草群落中底栖动物群落动态的初步研究[J].海洋湖沼通报,2006,(2):89-95.
[10] DAEHLER C C,STRONG D R.Variable reproductive output among clones of Spartina alterniflorainvading San Francisco Bay,California:the influence of herbivory,pollination,and establishment site[J].Am.J.Bot.,1994,81(3):307-313.
[11] GAO H,PENG X W,LI B,et al.Effects of the invasive plant Spartina alterniflora on insect diversity in Jiuduansha wetlands in the Yangtze River Estuary[J].Biodiversity Science,2006,14(5):400-409.高慧,彭莜葳,李博,等.互花米草入侵九段沙河口湿地对当地昆虫多样性的影响[J].生物多样性,2006,14(5):400-409.
[12] LU R K.Chemical analysis of agricultural soil[M].Beijing:China Agricultural Science and Technology Press,2000:1-163.鲁如坤.土壤农业化学分析方法[M].北京:中国农业科技出版社,2000:1-163.
[13] ZHONG H P,FAN J W,DU Z C,et al.The relationship between interspecific competition of ryegrass and quantification management indexes of pasture[J].Pratacultural Science,2004,21(5):30-37.钟华平,樊江文,杜占池,等.黑麦草种间竞争与草地数量化管理指数的关系[J].草业科学,2004,21(5):30-37.
[14] MAGUARAN A E.Ecological diversity and it measurement[M].NewJersey:Princeton university Press,1988:47-132.
[15] XU G W,ZHUO R Z.Preliminary studies of introduced Spartina(Spartina alterniflora Loisel)in China[J].Journal of Nanjing University,1985,40(2):212-225.徐国万,卓荣宗.我国引种互花米草(Spartina alterniflora Loisel)的初步研究(Ⅰ)[J].南京大学学报,1985,40(2):212-225.
[16] HE J,ZHAO C J,QING H,et al.Effect of soil-water condition on morphological plasticity of clonal plant Spartina alterniflora[J].Acta Ecologica Sinica,2009,29(7):3518-3524.何军,赵聪蛟,清华,等.土壤水分条件对克隆植物互花米草表型可塑性的影响[J].生态学报,2009,29(7):3518-3524.
[17] BRADLEY P M,MORRIS J T.Relative importance of ion exclusion,secretion,and accumulation in Spartina alterniflora Loisel[J].J.Exp.Bot.,1991,42(245):1525-1532.
[18] WANG Y P,ZHANG R S,GAO S.The dynamic response of the landscape of the salt marsh,tidal creek system[J].Chinese Science Bulletin,1998,43(21):2315-2320.汪亚平,张忍顺,高抒.论盐沼—潮沟系统的地貌动力响应[J].科学通报,1998,43(21):2315-2320.
[19] SUN S M.Monitoring of smooth cordgrass invasion by remote sensing in Sandy Bay,Fujian[J].Journal of Oceanography in Taiwan Strait,2005,24(2):223-228.
[20] SAYCE K,DUMBAULD B R,HIDY J.Seed dispersal in drift of Spartina alterniflora.[C]∥PATTEN K.Second International Spartina Conference.Pullman,WA.:Washington State University,1997:27-31.
[21] ZHANG R S,SHEN Y M,LU L Y,et al.Formation of Spartina alterniflora salt marshes on the coast of Jiangsu Province,China[J].Ecol.Eng.,2004,23:95-105.
(张 骞 编辑)
Competitive Ability of Two Propagules of Spartina alterniflora With Native Species in the Coastal Wetlands of North Jiangsu
CHEN Zheng-yong1,2,WANG Guo-xiang1,2,LIU Jin-e1,2,XU Wei-wei1,2,WANG Gang3
(1.College of Geography Science,Nanjing Normal University,Nanjing 210046,China;2.Jiangsu Key Laboratory of Environmental Change and Ecological Construction,Nanjing 210046,China;3.Anhui Normal University,Wuhu 241003,China)
The relatively competitive ability of S.alterniflora seedling and S.alterniflora tiller with Suaedaand reed was studied and to discuss the invasion trend of S.alterniflorainto the Suaeda salsaflat was discussed by selecting S.alterniflora patch in the Suaedaflat and dividing them into four sampling sites:periphery(A),outside(B),inside(C)and center(D).The results show that:(1)in site B the competitive ability of S.alterniflora seedling with Suaedais higher than that of S.alternifloratiller with Suaeda,while in site C the situation is just opposite.The diversity indices occurring in the sampling sites show an order of B>C>A>D.The significant values show an order of S.alternifloraseedling>Suaeda>reeds>S.alternifloratiller in site B,and S.alternifloratiller>reeds>S.alterniflora seedling>Suaedain site C.These indicate that S.alterniflorainvades into the Suaedaflat mainly by seedling and the population in the patch is kept stable mainly by tiller;(2)in site B,the competitive ability of reeds is comparable with that of S.alternifloraseedling,but higher than that of S.alternifloratiller.In site C,however,the competitive ability of S.alternifloratiller is higher than that of reeds,indicating that Suaeda might have a competing relation with native reed and could be replaced by both S.alterniflora and reed;and(3)The competitive ability of S.alternifloraseedling with Suaedahas a negative correlation to water content and total salinity of the flat soil(P<0.01)and a positive correlation to pH of the flat soil(P<0.01).However,the competitive ability of S.alternifloratiller with Suaedahas a positive correlation to water content and total salinity of the flat soil(P<0.05)and a negative correlation to pH of the flat soil(P<0.01).The water content and salinity of the soil in the Suaedaflat may possibly be favourable for S.alterniflorato settle down and to expand.
alien species;reproductive mode;expanding trend;interspecific competition;biological invasion
May 31,2011
Q948.12
A
1671-6647(2012)03-0380-10
2011-05-31
国家自然科学基金项目——互花米草对苏北盐沼沉积物有机碳库功能格局影响机理(40973053);江苏省高校自然科学重大基础研究项目——江苏省海滨湿地外来种互花米草扩张策略研究(08KJA180002)
陈正勇(1984-),男,江苏淮安人,硕士研究生,主要从事水环境生态学方面研究.E-mail:chenzhengyong111@163.com
*通讯作者,刘金娥(1973-),女,汉族,副教授,主要从事滨系统生态学方面研究.E-mail:liujine@njnu.edu.cn
