NaCl胁迫对四种荒漠植物种子萌发的影响
2012-08-20杨景宁王彦荣
杨景宁,王彦荣
(草地农业生态系统国家重点实验室 兰州大学草地农业科技学院,甘肃 兰州730020)
我国的干旱地区大多为生态脆弱区,同时又为盐碱区[1]。探讨这些地区的植物种子萌发问题,对其植被的恢复与重建具有至关重要的意义。干旱和盐分是影响这些地区植物种子萌发的两大关键性因素,本研究侧重于探讨盐分这一重要因素。
盐分对种子萌发的影响一般归结为渗透效应与离子效应[2,3],种子耐盐性的实质是种子萌发过程中对盐分造成的渗透效应与离子效应的综合适应,而生态因子(如盐分类型和温度等)则通过改变渗透效应与离子效应的大小来影响种子耐盐性[4]。总体而言,种子萌发是各种生态因子相互作用的产物,但萌发对生态因子的响应程度,因物种和物种生态型的不同而异。
本研究选择内蒙古阿拉善荒漠区的4种优势建群植物种:梭梭(Haloxylon ammodendron)、红砂(Reaumuria soongorica)、驼绒藜(Ceratoides latens)和碱蓬(Suaeda glauca)为材料。在气候干旱,土地盐渍化严重[5]的阿拉善荒漠,这4种植物广泛分布,它们是良好的固沙植物,同时也是干旱荒漠区家畜在干旱季节的主要采食对象。
关于这4种植物的萌发生态学研究,已有资料涉及温度、光照、盐分和贮藏条件对梭梭种子萌发的影响[6-8];温度、光照、干旱和播深对红砂种子萌发的影响[9,10];温度、光照、贮藏和发芽床等因素对驼绒藜种子萌发的影响[11-13];光照、温度和盐分对碱蓬种子萌发的影响[14-16]。
由此可见,有关这4种荒漠植物种子萌发对盐胁迫响应的综合比较研究还比较少,而经NaCl浸种预处理后,种子在适宜条件下的萌发研究则鲜见报道。本研究从生理生态学的角度系统比较了4种荒漠植物种子萌发对盐分胁迫(NaCl)的响应,以及经NaCl浸种预处理后,种子在适宜条件下的萌发表现,以期为促进我国荒漠植物种子的抗逆性研究,为开发利用荒漠植物种质资源和进行植被的恢复与重建提供参考和借鉴。
1 材料与方法
1.1 研究材料
供试4种荒漠植物梭梭(藜科,灌木)、红砂(柽柳科,小灌木)、驼绒藜(藜科,半灌木)和碱蓬(藜科,草本)种子于2005年10月下旬采收于内蒙古阿拉善左旗的荒漠草原,每种植物种子均包含至少100株的混合种子,经测定千粒重分别为3.3,1.3,3.9和2.1g。种子风干后,置于-5℃低温保存。试验于2006年6月-12月在兰州大学、农业部牧草与草坪草种子质量监督检验测试中心(兰州)进行。
1.2 研究方法
1.2.1 发芽基本条件 发芽采用培养皿纸上法,参照《国际种子检验规程》[17],文献报道[9,13,18]和预备试验,确定25℃为4种供试植物种子的适宜萌发温度,在培养箱无光照条件下发芽。各处理均为4次重复,每重复50粒种子,发芽期为15d。
1.2.2 NaCl盐胁迫处理 NaCl溶液浓度梯度设置:0(对照),0.05,0.1,0.2,0.3,0.4,0.5,0.6,0.7,0.8,0.9,1.0,1.2,1.4,1.6和1.8mol/L的NaCl溶液共16个处理。各处理培养皿中放入2张用7mL NaCl溶液浸湿的滤纸,摆入种子。每天以称重法加入蒸馏水以保持溶液的浓度恒定,逐日统计发芽数,计算发芽率和发芽指数,7 d测定幼苗的胚芽(包括胚轴与顶芽)和初生根长度。
1.2.3 NaCl浸种预处理后种子的萌发 在4种荒漠植物种子萌发的盐度阈值(梭梭:1.8mol/L;红砂:0.6 mol/L,驼绒藜:0.9mol/L;碱蓬:0.5mol/L)下,浸种1,3,5,8,10,15,20,25,30,35,40,45和50d,至不能萌发为止。然后用不同浸种天数的种子在适宜条件下进行复萌实验,以未经浸种处理的种子至为对照。
1.3 数据分析方法
所有数据均用Excel统计并制图。应用SPSS 11.0统计分析软件对不同处理下的发芽率、发芽指数、初生根长和胚芽长等各项指标进行差异显著性分析。
2 结果与分析
2.1 盐胁迫对4种荒漠植物种子发芽率的影响
不同浓度的NaCl对4种荒漠植物种子的发芽率具有抑制作用,且萌发抑制程度一般随盐溶液浓度的升高而增加,但不同植物对盐胁迫的响应程度不同(图1)。梭梭种子的发芽率在0~0.8mol/L变化不大,之后随NaCl浓度的增加发芽率呈下降趋势,但其发芽率始终显著高于其他3种植物。1mol/L盐度下,梭梭种子仍有88%的萌发率,而其他3种植物的萌发率已经为零。驼绒藜和碱蓬种子的发芽率随着NaCl浓度的升高先增加(驼绒藜0~0.3mol/L,碱蓬种子0~0.1mol/L)后降低。当NaCl浓度达到0.4mol/L时,驼绒藜种子的发芽率显著高于碱蓬和红砂;当NaCl浓度大于0.4mol/L时,红砂种子的发芽率开始高于碱蓬,但二者的发芽率没有显著性差异。梭梭、红砂、驼绒藜和碱蓬种子萌发的耐盐极限值分别为:1.8,0.6,0.9和0.5mol/L,从高到低应依次为:梭梭>驼绒藜>红砂>碱蓬。
2.2 盐胁迫对4种荒漠植物种子发芽指数的影响
梭梭种子的发芽指数在0~0.4mol/L变化不大,之后随NaCl浓度的增加发芽指数呈下降趋势,但其发芽指数始终显著高于其他3种植物(图2)。碱蓬和驼绒藜种子的发芽指数均呈先增加后降低的趋势。当NaCl浓度小于0.3而大于等于0.1mol/L时,4种荒漠植物种子的发芽指数从高到低的显著性序列为:梭梭>碱蓬>驼绒藜>红砂;而驼绒藜与碱蓬种子的发芽指数在NaCl浓度为0.3mol/L时无显著性差异,和其他3种植物相比,红砂种子的发芽指数始终最小。
2.3 盐胁迫对4种荒漠植物初生根生长的影响
不同浓度的NaCl对4种荒漠植物的初生根生长具有不同程度的抑制作用,且抑制程度随盐溶液浓度的升高而增加(图3)。对照条件下,碱蓬的初生根长显著高于其他3种植物,而红砂和驼绒藜的初生根长却没有显著性差异,且二者均显著高于梭梭;当NaCl浓度小于0.2mol/L时,4种荒漠植物的初生根长存在显著性差异;在0~0.4mol/L范围内碱蓬的初生根长始终显著高于其他3种植物,4种荒漠植物的初生根长排序为:碱蓬>红砂>驼绒藜>梭梭;在NaCl浓度为0.3mol/L时,驼绒藜和红砂、梭梭和红砂的初生根长之间没有显著性差异;在NaCl浓度为0.4mol/L时,驼绒藜和梭梭的初生根长显著高于红砂,而它们二者之间却没有显著性差异。0.9 mol/L的NaCl浓度下,梭梭仍能生长,而其他3种植物的初生根生长已经完全被抑制。
2.4 NaCl浸种预处理后4种荒漠植物种子的萌发

图1 4种荒漠植物种子在不同浓度NaCl溶液中的发芽率Fig.1 Germination percentage of four desert plant seeds at different concentration of NaCl solution

图2 4种荒漠植物种子在不同浓度NaCl溶液中的发芽指数Fig.2 Germination index of four desert plant seeds at different concentration of NaCl solution

图3 4种荒漠植物种子在不同浓度NaCl溶液中的初生根长度Fig.3 Radical length of four desert plant seeds at different concentration of NaCl solution
NaCl浸种预处理后4种荒漠植物种子的萌发结果显示,4种荒漠植物种子对NaCl胁迫响应不同,解除NaCl胁迫后,红砂种子的发芽率(第1天)和发芽指数(第1到25天)均呈先增加后降低的趋势,而其他3种植物种子的发芽率和发芽指数均呈下降趋势(表1和2)。与对照相比,解除NaCl胁迫后,梭梭种子的发芽率在NaCl浸种前3d,发芽指数在NaCl浸种前5d没有显著性差异,之后开始显著下降;驼绒藜和碱蓬种子的发芽率、发芽指数均在NaCl浸种1d后其就开始显著下降,且NaCl浸种1至3d,碱蓬种子的发芽率没有显著性差异;在NaCl浸种约10d时,解除胁迫后,碱蓬种子的萌发完全被抑制,而此时梭梭种子的复萌率显著高于红砂,红砂种子的复萌率显著高于驼绒藜,复萌序列为梭梭>红砂>驼绒藜>碱蓬,之后红砂的复萌率显著高于其他植物;梭梭、红砂、驼绒藜和碱蓬种子完全被抑制萌发的浸种天数分别约为:25,50,20和10d。
2.5 NaCl浸种预处理后4种荒漠植物的初生根生长
解除NaCl胁迫后,除碱蓬的初生根随着盐胁迫天数的延长呈降低趋势外,其他3种荒漠植物的初生根长均随着盐胁迫天数的延长呈现出先增加(梭梭和红砂第1至8天,驼绒藜第1天)后降低的趋势(表3)。和对照相比,在NaCl浸种1至3d后梭梭的初生根长显著增加,分别是对照的1.8和1.7倍,在NaCl浸种8至10d时,其初生根和对照没有显著性差异,之后呈显著降低的趋势;在NaCl浸种第25天后,红砂的初生根仍能继续生长,而其他3种植物的初生根已经被完全抑制生长;在NaCl浸种1d时驼绒藜的初生根和对照之间没有显著性差异,之后呈显著下降的趋势;碱蓬的初生根在NaCl浸种1d时呈下降趋势,浸种3d时和对照之间没有显著性差异。
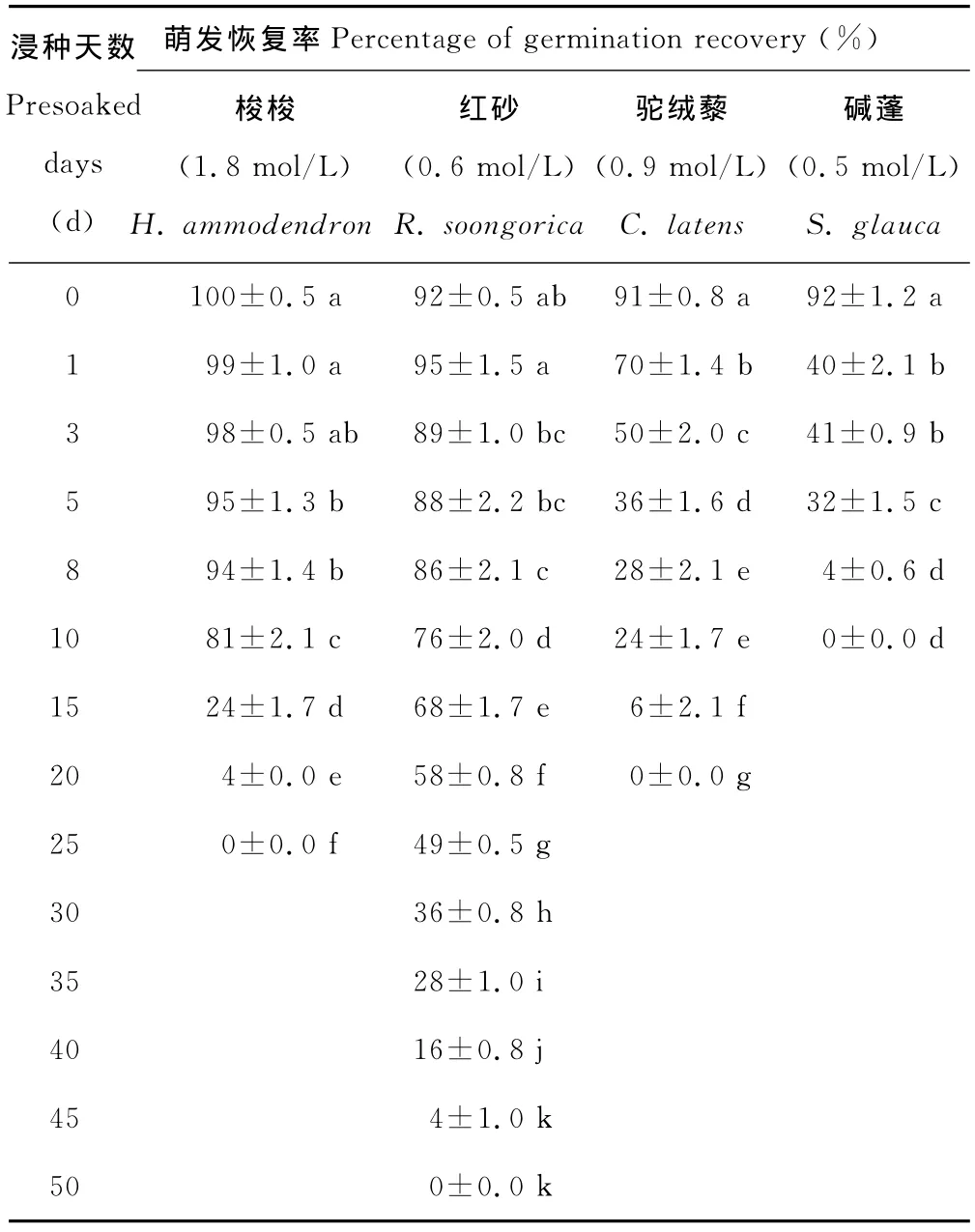
表1 4种荒漠植物种子经NaCl浸种预处理后的萌发恢复率Table 1 Percentage of germination recovery of four desert plant seeds after presoaking with NaCl solution
3 讨论
3.1 盐胁迫对4种荒漠植物种子萌发的影响
总体来看,不同浓度的NaCl对4种荒漠植物种子萌发均具有一定的抑制作用(图1和2),许多研究都有类似的报道[14,19,20]。盐胁迫下,梭梭种子的发芽率和发芽指数始终显著高于其他3种植物,在1mol/L盐浓度下,梭梭种子仍有88%的萌发率和10.33的发芽指数,而其他3种植物种子的萌发率和发芽指数已经为零,这表明梭梭种子不但发芽整齐而且耐盐性很强。当NaCl浓度较低时,驼绒藜种子的发芽率、碱蓬种子的发芽率和发芽指数都有增加的趋势,说明轻度的NaCl胁迫能促进这两种植物种子的萌发,并能提高碱蓬种子的发芽速率和整齐度。可能是由于种子从盐溶液中吸收无机盐离子,增加了细胞溶液浓度,降低了细胞水势,从而增加了种子吸水的能力,进而提高了其萌发率。但是,盐分对梭梭种子的萌发却没有促进作用,这与黄振英等[8]的研究一致。红砂种子在盐胁迫下表现出独特的反应机制,发芽指数低、萌发持续而分散,既说明盐分对红砂种子的萌发具有一定的抑制作用,又表明红砂种子可能对盐分有一定的适应性。
盐分对4种荒漠植物的初生根生长具有不同程度的抑制作用,抑制程度随盐溶液浓度的升高而增加(图3)。这与王玉祥等[21]、景艳霞和袁庆华[22]对苜蓿的研究一致。0.9mol/L的NaCl浓度下,梭梭的初生根仍能生长,而其他3种植物的初生根的生长已经完全被抑制,说明梭梭幼苗的抗盐性很强,其对盐渍生境的适应性较强。在0~0.4mol/L范围内碱蓬的初生根长始终显著高于其他3种植物,说明碱蓬能在低盐环境下较好地生长,这可能也体现了一年生草本植物对生境的特殊适应机制。

表2 4种荒漠植物种子经NaCl浸种预处理后的发芽指数Table 2 Germination index of four desert plant seeds after presoaking with NaCl solution

表3 4种荒漠植物种子经NaCl浸种预处理后的初生根长度Table 3 Radical length of four desert plant seeds after presoaking with NaCl solution
藜科荒漠植物的种子(如梭梭)在低盐溶液中的萌发率和在无盐条件下的情况差别不太大(图1),这和Katembe等[23]报道的结果一致。随着盐溶液浓度的升高,萌发过程逐渐受抑制[8,19,24],但是各种盐生植物对盐度的耐受阈限不同,相同盐浓度下的萌发率也不同[25,26],红海蓬子(Salicorniarubra)[27]和盐生草(Halogeton glomeratus)[28]的种子在1 000mmol/L NaCl溶液中仍能萌发,而盐爪爪和盐穗木种子分别在540和749mmol/L的盐浓度下就停止了萌发。本研究中梭梭、红砂、驼绒藜和碱蓬种子萌发的耐盐极限值分别为1.8,0.6,0.9和0.5mol/L,表明梭梭种子发芽耐盐性最强,其次为驼绒藜,而碱蓬的耐盐性最弱。
盐胁迫下,植物种子的萌发生长与渗透胁迫和离子毒害有关,而这两个因素与种子萌发的生理学特征密切相关[19,25,29-31]。曾幼玲等[20]研究认为盐生植物中起主要抑制作用的是渗透胁迫而不是离子毒害,而张万钧等[30]认为影响盐生植物种子萌发的因素不仅仅是渗透胁迫,NaCl可能对在种子萌发中起关键作用的一些酶的活动有抑制作用。另外,植物在种子萌发期的耐盐性表现与发育阶段不同,可能是耐高盐植物在种子阶段进入了由渗透胁迫引起的休眠状态,在一定程度上避免了高盐的毒害甚至致死作用,当有一定量的雨水时,土壤盐度降低,促使一些植物种子迅速吸水、萌发、生长,从而完成生活史[32-36]。植物种子萌发是一个极为复杂的生理过程,耐盐能力的大小是多种代谢的综合表现[37]。盐害机理是当今研究的热点,本实验只对4种荒漠植物种子萌发和初生生长阶段的盐害机理进行了初步探讨,但盐分对植物不同生长发育阶段或不同生理过程的影响,有待于进一步探讨研究。
3.2 NaCl浸种预处理后4种荒漠植物种子的萌发
在高盐环境下,种子能否保持活力及幼苗能否继续生长是植物存活的关键[19],当胁迫条件减轻时,大多数盐生植物显示出明显的复萌状态,表明它们可能比正常生长的植物更耐盐[38]。随着浸种天数的延长,红砂种子的发芽率和发芽指数呈现出先增加后降低的趋势,而其他3种植物种子的发芽率和发芽指数均呈下降趋势(表1和2),说明短时间的NaCl浸种对红砂种子的萌发有一定的促进作用,可显著提高红砂种子的发芽速度和整齐度,但浸种时间越长,种子受盐胁迫的影响越大。NaCl浸种25d时,解除浸种胁迫后的红砂种子仍有49%萌发率,而其他3种植物种子的萌发已经完全被抑制,说明红砂种子对盐分有特殊的适应机制。这些结果同时表明,盐浸种后碱蓬等藜科植物种子受到伤害较大,萌发恢复力弱,而柽柳科红砂的种子受到的伤害较小,萌发恢复力也较强。
梭梭、红砂和驼绒藜的初生根长均随着浸种天数的增加呈现出先增加后降低的趋势(表3),说明短时间的NaCl浸种预处理对这3种植物的初生根生长有一定的促进作用,尤其是梭梭,NaCl浸种1到3d后其初生根长势良好。与此相反,NaCl浸种抑制了碱蓬的初生根生长。NaCl浸种25d后,红砂的初生根仍能生长,其他3种植物的初生根生长已经被完全抑制,说明盐溶液浸种有益于红砂幼苗的生长,这可能是红砂长期适应自然环境的结果。这些结果同时说明,NaCl浸种处理后草本植物碱蓬的初生根生长始终受到抑制,但却在一定程度上促进了其他3种灌木植物梭梭、红砂和驼绒藜的初生根生长。长时间来看,NaCl浸种预处理还是对4种荒漠植物的初生根生长具有抑制作用。
种子恢复后的活力是决定能否建植成功的因素之一,盐浸种后,种子可以恢复萌发,但其初生根和胚芽有无较强的生命力也很重要。本研究结果表明,梭梭、红砂和驼绒藜种子经短时间高浓度的盐溶液浸种预处理后,其初生根生长较好,这种对盐分的适应性说明其适于盐碱环境的生存,这对于荒漠地区植被的恢复与重建具有重要意义。
[1] 徐树建.我国西北地区生态恢复研究[J].地理学与国土研究,2002,18(2):80-84.
[2] Levitt J.Responses of Plants to Environmental Stress(2nd ed.)[M].New York:Academic Press,1980:365-434.
[3] Ungar I A.Halophyte seed germination[J].Botanical Review,1978,44:233-264.
[4] 阎顺国,沈禹颖.生态因子对碱茅种子萌发期耐盐性影响的数量分析[J].植物生态学报,1996,20(5):414-422.
[5] 杜方红,黄文浩.阿拉善地区生态环境问题及探讨[J].内蒙古环境保护,2005,17(3):5-9.
[6] Tobe K,Li X,Omasa K.Effects of sodium chloride on seed germination and growth of two Chinese desert shrubs,Haloxylon ammodendronand H.persicum (Chenopodiaceae)[J].Australian Journal of Botany,2000,48:455-460.
[7] Wang X Y,Wei Y,Yan Ch,et al.Germination response of two species of Haloxylonto temperature and salinity[J].Arid Zone Research,2004,21(S):58-63.
[8] 黄振英,张新时,Yitchak Gutterman,等.光照、温度和盐分对梭梭种子萌发的影响[J].植物生理学报,2001,27(3):275-280.
[9] 曾彦军,王彦荣,张宝林,等.红砂和猫头刺种子生态适应性的研究[J].草业学报,2000,9(3):36-42.
[10] 曾彦军,王彦荣,庄光辉,等.红砂和霸王种子萌发对干旱与播深条件的响应[J].生态学报,2004,24(8):1629-1634.
[11] 李青丰,易津.驼绒藜属植物种子萌发检验标准及幼苗发育特性的研究[J].内蒙古草业,1994,(1-2):47-49.
[12] 易津,曹自成,乌仁其木格.几种不同贮藏条件对华北驼绒藜种子寿命和活力的影响[J].内蒙古农牧学院学报,1994,15(1):18-22.
[13] 王学敏,易津.驼绒藜属植物种子萌发条件及其生理特性的研究[J].草地学报,2003,11(2):95-102.
[14] 段德玉,刘小京,冯凤莲,等.不同盐分胁迫对盐地碱蓬种子萌发的效应[J].中国农学通报,2003,19(6):168-172.
[15] 李昀,沈禹颖,阎顺国.NaCl胁迫下5种牧草种子萌发的比较研究[J].草业科学,1997,14(2):50-53.
[16] 王雷,田长彦,张道远,等.光照、温度和盐分对囊果碱蓬种子萌发的影响[J].干旱区地理,2005,28(5):670-674.
[17] 国际种子检验学会(ISTA).国际种子检验规程[S].北京:技术标准出版社,1980:145-183.
[18] 张树新,邹受益,杨美霞.梭梭种子发芽特性实验研究[J].内蒙古林学院学报,1995,17(2):56-63.
[19] 沈禹颖,王锁民,陈亚明.盐胁迫对牧草种子萌发及其恢复的影响[J].草业学报,1999,8(3):54-60.
[20] 曾幼玲,蔡忠贞,马纪,等.盐分和水分胁迫对两种盐生植物盐爪爪和盐穗木种子萌发的影响[J].生态学杂志,2006,25(9):1014-1018.
[21] 王玉祥,陈爱萍,张博.不同苜蓿品种耐盐性初探[J].草业科学,2010,27(3):102-106.
[22] 景艳霞,袁庆华.NaCl胁迫对苜蓿幼苗生长及不同器官中盐离子分布的影响[J].草业学报,2011,20(2):134-139.
[23] Katembe W J,Ungar I A,John P M.Effect of salinity on germination and seedling growth of two Atriplexspecies[J].An-nals of Botany,1998,82:167-175.
[24] Khan M A,Ungar I A.Effects of light,salinity,and thermoperiod on the seed germination of halophytes[J].Canadian Journal of Botany,1997,75(5):835-841.
[25] 李海云,赵可夫,王秀峰.盐对盐生植物种子萌发的抑制[J].山东农业大学学报(自然科学版),2002,33(2):170-173.
[26] Khan M A,Gul B,Weber D J.Seed germination in the Great Basin halophyte Salsola iberica[J].Canadian Journal of Botany,2002,80:650-655.
[27] Khan M A,Gul B,Weber D J.Germination responses of Salicornia rubra to temperature and salinity[J].Journal of Arid Environments,2000,45:207-214.
[28] Khan M A,Gul B,Weber D J.Seed germination characteristics of Halogetonglomeratus[J].Canadian Journal of Botany,2001,79:1189-1194.
[29] 侯旭光,赵可夫.非盐生植物棉花和盐生植物灰绿藜的盐害机理[J].山东大学学报(自然科学版),1999,34(2):230-235.
[30] 张万钧,王斗天,范海,等.盐生植物种子萌发的特点及其生理基础[J].应用与环境生物学报,2001,7(2):117-121.
[31] Wang B,Luttge U,Ratajczak R.Effects of salt treatment and osmotic stress on V-ATPase and V-PPase in leaves of the halophyte Suaeda salsa[J].Journal of Experimental Botany,2001,52:2355-2365.
[32] 赵可夫,冯立田,范海.盐生植物种子的休眠、休眠解除及萌发的特点[J].植物学通报,1999,16(6):677-685.
[33] Ungar I A.Population ecology of halophyte seeds[J].Botanical Review,1987,53:301-344.
[34] Ungar I A.Seed germination and seed-bank ecology in halophytes[A].In:Kigel J,Galili G.Seed Development and Germination[M].NewYork:Marcel Dekker,1995:599-628.
[35] Tobe K,Li X,Omasa K.Seed germination and radicle growth of a halophyte,Kalidium caspicum[J].Annals of Botany,2000,85:391-396.
[36] 秦峰梅,张红香,武祎,等.盐胁迫对黄花苜蓿发芽及幼苗生长的影响[J].草业学报,2010,19(4):71-78.
[37] 李源,刘贵波,高洪文,等.紫花苜蓿种质耐盐性综合评价及盐胁迫下的生理反应[J].草业学报,2010,19(4):79-86.
[38] Pujol J A,Calvo J F,Ramirez D L.Recovery of germination from different osmotic conditions by four halophytes from Southeastern Spain[J].Annals of Botany,2000,85:279-286.
