高原地区汉、藏、回族人群类风湿关节炎与肽酰基精氨酸脱亚氨酶4基因多态性的相关性
2012-08-02杨发满张培莉汪元浚青海大学附属医院老年2科青海西宁810001
刘 冀 杨发满 张培莉 李 蓉 汪元浚 (青海大学附属医院老年2科,青海 西宁 810001)
本研究对90例类风湿关节炎(RA)患者(汉族、回族、藏族)和90名健康对照肽酰基精氨酸脱亚氨酶4(PADI4)-92、94和104的单核苷酸多态性(SNP)进行分析,以探讨其与RA易感性的关系。
1 资料与方法
1.1 临床资料 收集2010年1月至2011年12月,我院风湿科住院及门诊RA汉族患者(海拔2 200~2 600 m)30例,玉树地区住院及门诊RA藏族(海拔3 800~4 200 m)患者30例,大通塔尔住院及门诊RA回族患者(海拔2 700 m)30例,诊断均符合美国风湿病学会1987年RA分类诊断标准,并排除其他自身免疫性疾病;其中男22例,女68例,平均年龄(50±14)岁;健康对照组90名,分别为各地区各民族健康体检者,其中男15名,女75名,平均年龄(49±12)岁。研究对象均无血缘关系,各组年龄及性别等资料差异无统计学意义(P>0.05)。
1.2 基因组DNA提取 采集受检者静脉血5 ml,乙二胺四乙酸二钠(EDTA-Na:)抗凝;采用AxyPrep-96全血基因组DNA试剂盒提取白细胞基因组DNA,0.8%琼脂糖凝胶电泳检测其完整性,-80℃保存备用,AxyPrep-96全血基因组DNA试剂盒购自AXYGEN公司。
1.3 PCR反应 在MJ Research PTC-250梯度PCR仪上进行。PCR反应结束后,取PCR反应产物5 μl加1 μl加样缓冲液在1.5%琼脂糖凝胶上电泳。电压50 V,电泳时间30~40 min。紫外光下观察电泳结果,使用凝胶成像系统拍照记录。引物序列见表1。以ABI PRISM 3730 DNA测序仪测定LDR产物序列长度后经Genemapper分析得出SNP基因型分型结果。

表1 引物序列
1.4 其他实验室指标测定 抗环瓜氨酸肽(CCP)抗体采用ELISA法(博尔迈医学实验诊断有限公司),红细胞沉降率(ESR)采用魏氏法检测。
1.5 RA活动性评分标准 RA疾病活动关节评分28(DAS28)以28个关节计分DAS28≥5.1为疾病高度活动;3.2≤DAS28<5.1为疾病中度活动;2.6≤DAS28<3.2为疾病低度活动;DAS28<2.6为病情缓解。
1.6 健康评定问卷(HAQ)评分 采用HAQ问卷对RA患者生存质量进行评分。HAQ残疾指数计算方法:在过去的1个月内从事以下活动的评价,①自己穿衣服,包括系鞋带和纽扣;②上床、下床;③端一满杯水送到嘴边;④在室外的平地上行走;⑤自己洗澡,且擦干身体;⑥蹲下,拾起地上的衣服;⑦上、下车。每项得分如下:0=无困难,1=有些困难,2=很困难,3=不能进行。把各项分相加即为总得分。
1.7 统计学方法 应用SPSS17.0统计软件进行分析,用r吻合度检验分析患者基因型分布是否符合Hardy-Weinberg定律,各基因型及等位基因频率比较采用χ2检验,各基因型与临床资料相关性分析采用方差分析、秩和检验。
2 结果
2.1 Hardy-Weinberg平衡分析 经χ2检验,RA组和健康对照组 PADI4-94、PADI4-104、PADI4-92 位点实际基因分布与 Hardy-Weinberg平衡状态下的理论分布差异无统计学意义(χ2=1.760,P=0.184; χ2=1.191,P=0.275; χ2=1.679,P=0.192),表明均符合该平衡法则,具有群体代表性。
2.2 不同民族RA组与对照组基因型及其频率 PADI4-92、PADI4-94及PADI4-104位点存在A和 G、C三种等位基因,A/A、C/C、G/G、A/G,G/C 五种基因型、汉族、回族、藏族 RA 组、健康对照组 PADI4-94 PADI4-104的A/A、G/G和 A/G基因型频率,组间差异无统计学意义(χ2=1.105,P=0.949);(χ2=1.105,P=0.949);(χ2=0.657,P=0.720)。(χ2=0.617,P=0.734);(χ2=0.617,P=0.734);(χ2=1.198,P=0.549)。汉族、回族、藏族RA组、健康组PADI4-92的C/C、G/G和G/C基因型频率组间差异无统计(χ2=1.105,P=0.949)(χ2=1.105,P=0.949)(χ2=0.234,P=0.889)。见表 2。
2.3 RA 组 PADI4-94 及 PADI4-104、PADI4-92 各基因型间临床指标的比较 ESR、DAS28评分、HAQ生活质量评分及抗CCP 抗体在 RA 组 PADI4-94 及 PADI4-104、PADI4-92 各基因型间差异无统计学意义(P>0.05)。
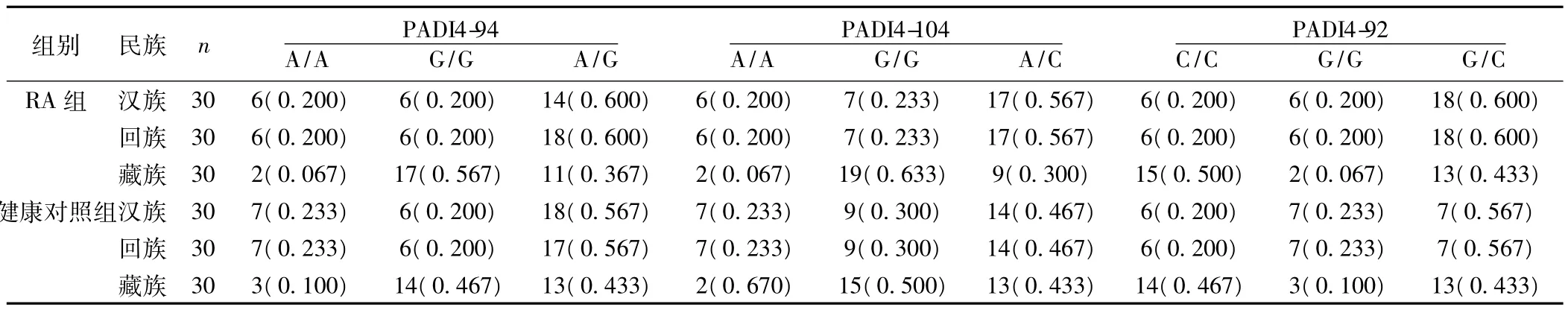
表2 RA组与健康对照组PADI4-94、PADI4-104、PADI4-92基因型及其频率〔n(频率)〕
3 讨论
PADI基因是1981年Fujisaki等首先从新生大鼠表皮提取物中分离得到的一种基因。目前,在人体组织中共发现5种PADI酶(即 PADll,2、3、4、6),其中,PADI4 与 RA 关系最为密切。人类PADI4基因定位于染色体1p36上的NT.034367.1区域。PADI是一种翻译后修饰酶,可以催化抗原蛋白多肽进行瓜氨酸化反应,并在钙离子存在的情况下把精氨酸残基转化成瓜氨酸残基,形成瓜氨酸肽,打破机体的免疫耐受而诱发RA的发生。由于瓜氨酸肽的形成有赖于PADI的存在。因此,PADI基因是RA发病的重要因素。研究证实,PADI4在人细胞信号传导途径发挥调节作用,包括细胞分化、细胞凋亡和基因转录等〔1〕。在整个PADI4基因上,从5′非翻译区的启动子序列到3′非翻译区,目前已发现多个 SNP位点,PADI4-89(外显子2,163G→A)、PADI4-90(外显子 2,245G→C)、PADI4_92(外显子3,335G→C)、PADI4-104(外显子 4,349T→C)以及内含子区域PADI4_94(内含子3,28017T→C)的SNPs是研究的热点。X射线分析表明这些氨基酸置换发生在肽链N端,远离蛋白活性部位,并不会直接影响PADI4的催化功能,也不影响二聚体的连接,目前尚不清楚这些突变是否影响到酶与核受体或靶蛋白的相互作用〔2〕。故深入探讨PADI4与RA易感性有助于进一步阐明RA的病理及发病机制。
RA是一种具有遗传倾向的多基因疾病,在遗传因素中,人类白细胞抗原(HLA)所起的作用也只有30% ~50%,其余50%-70%的HLA影响RA易感性的基因分布至今未明。Suzuki等〔3〕采用SNP方法对日本人进行病例对照研究发现,PADI4基因与RA的易感性相关。该研究通过分析PADI4基因中的17 个 SNPs,发现其中的 8 个 SNPs(PADI4-92、PADI4-94、PADI4-104、PADI4-95、PADI4-97、adi4_99、PADI4-100、PADI4-101) 与RA有很强的相关性。其中PADI4-94与RA的联系尤为显著。日本、韩国研究也证实了RA与PADI4的相关性。Plenge等〔4〕在北美和瑞典人群中对RA关联基因进行了研究,发现PADI4-94是RA的易感基因,可见PADI4是RA易感基因。本研究结果显示 PADI4-94 及 PADI4-104、PADI4-92 基因多态性与 RA 易感性无关,与我国史恒星等〔5〕及西班牙 Martinez等〔6〕研究结果一致。此外,在钱龙等〔7〕研究证实,PADI4-92基因多态性与RA无明显相关性也与本研究结果符合。随后,Barton等〔8〕利用连锁与关联的基本策略分析方法对英国人群中PADI4基因进行分析,PADI4基因的SNPs或单体型均与RA发病无关。同时,在对德国人和西班牙人的研究中也发现PADI4基因多态性与RA无相关性。PADI4与RA的关联性可能取决于种群之间的遗传异质性。
Vossenaar等〔9〕报道了蛋白质瓜氨酸化是在PAD酶的作用下精氨酸残基发生翻译后修饰形成瓜氨酸残基的过程。通常情况下,PAD酶以非活性形式存在于细胞内,当细胞死亡后,胞膜的完整性缺失,胞内PAD酶外流,在细胞外足够高的Ca2+浓度的环境下,PAD酶被激活并导致细胞外蛋白质的瓜氨酸化。患者的炎症关节滑膜组织中出现瓜氨酸化蛋白并非RA所特有,瓜氨酸化蛋白也可以在如骨关节炎、反应性关节炎等其他患者的炎症关节滑膜组织中检出,蛋白质瓜氨酸化是与关节炎症相关的标志而非RA特异性标志,而瓜氨酸化蛋白驱动的瓜氨酸化蛋白特异性B细胞的成熟以及由此产生的抗CCP抗体才是RA所特有的,抗CCP抗体促使关节炎症持续存在并转化为慢性RA或重症RA。Cha等〔10〕报道,PADI4可能在RA早期(≤34个月)发挥作用,并影响抗CCP抗体的产生。但大多欧美国家研究均表明,PADI4各基因型与抗CCP抗体无相关性,可见PADI4参与了蛋白质的瓜氨酸化,参与了RA发病,但在不同种族中可能作用不同。本研究显示,RA患者血清中PADI4水平与抗CCP抗体无相关性,与大多数欧美国家PADI4基因层面的研究结果一致。
可见,PADI4可能参与了RA发病,间接参与RA抗体的形成及关节破坏进展。本研究中PADI4基因与抗CCP抗体形成无相关性,可能与样本量小及病例选择偏倚有关。除PADI4-94和 PADI4-104、PADI4-92 三型,抗 CCP 抗体形成可能与 PADI4基因其他亚型相关,这有待于大样本多亚型研究。
1 Arita K,Shimizu T,Hashimoto H,et al.Structural basis for histone N-terminal recognition by human peptidylarginine deiminase 4〔J〕.Proc Natl Acad Sci USA,2006;103(14):5291-6.
2 Arita K,Hashimoto H,Shimizu T,et al.Structural basis for Ca(2+)induced activation of human PAD 4〔J〕.Nat Struct Mol Biol,2004;11(8):777-83.
3 Suzuki M,Miyagi J,Kuribayashi M,et al.Evaluation of allele frequencies in the PADI4 gene and anti-cyclic eitrullinated peptide antibodies of patients with rheumatoid arthritis in a Japanese population〔J〕.Ann Rheum Dis,2006;65(10):1399-400.
4 Plenge RM,Padyukov L,Remmers EF,et al.Replication of putative candidate-gene association with rheumatoid arthritis in > 4000 samples from North America and Sweden:association of susceptibility with PTPN22,CTLA4,and PADI4〔J〕.Am J Hum Genct,2005;77(6):1044-60.
5 史恒星,钱 龙,李向培,等.肽酰基精氨酸脱亚氨酶-4基因多态性与类风湿关节炎的相关性〔J〕.中华风湿病学杂志,2010;14(5):336-9.
6 Martinez A,Valdivia A,Pascual-Salcedo D,et al.PADI4 polymorphisms are not associated with rheumatoid arthritis in the Spanish population〔J〕.Rheumatology(Oxford),2005;44(10):1263-6.
7 钱 龙,史恒星,李向培,等.类风湿关节炎患者血清肽酰基精氨酸脱亚氨酶4水平及其意义〔J〕.中华内科杂志,2011;50(2):107-10.
8 Barton A,Bowes J,Eyre S,et al.A functional haplotype of the PADI4 gene associated with rheumatoid arthritisin a Japanese population is not associated in Aunited Kingdom population〔J〕.Arthritis Rheum,2004;50(4):1117-21.
9 Vossenaar ER,van Venrooij WJ.Citrullinated proteins:sparks that may ignite the fire in rheumatoid arthritis〔J〕.Arthritis Res Ther,2004;6(3):107-11.
10 Cha S,Choi CB,Han TU,et al.Association of anti-cyclic citrullinated peptide antibody levels with PADI4 haplotypes in early rheumatoid arthritis and with shared epitope alleles in very late rheumatoid arthritis〔J〕.Arthritis Rheum,2007;56(5):1454-63.
