罗非鱼营养需求研究进展
2012-07-12中国水产科学研究院淡水渔业研究中心
中国水产科学研究院淡水渔业研究中心
农业部淡水渔业和种质资源利用重点实验室 乐贻荣 杨 弘
罗非鱼是重要的淡水养殖鱼类之一。为配合罗非鱼的规模化养殖,促进罗非鱼产业的快速发展,大量学者对罗非鱼的营养需求进行了研究。本文就不同品系罗非鱼,包括奥利亚罗非鱼、尼罗罗非鱼及奥尼罗非鱼等的营养需求研究作一综述。
1 蛋白质和氨基酸
1.1 蛋白质 蛋白质是生命的物质基础,是所有生物体的重要组成成分,是决定鱼类生长的最关键的营养物质之一。不同品系罗非鱼对蛋白质需要量见表1。由表1可知,不同品系罗非鱼之间对蛋白质需求量存在一定差异,而且同一品系罗非鱼在不同的生长阶段对蛋白质需求量也是不同的。由于奥尼罗非鱼和尼罗罗非鱼是我国目前最主要的养殖品种,因此进一步完善其各生长阶段,如仔稚和养成阶段的蛋白质需求量,对于指导实际生产具有重要意义。
1.2 氨基酸 鱼类对蛋白质的需要实际是对氨基酸的需要,尤其是对必需氨基酸的需要。罗非鱼对必需氨基酸需要量见表2。由于不同品系的罗非鱼对蛋白质的需求量存在较大差异,而且目前罗非鱼对必需氨基酸的需求量研究结果大多是较小规格的罗非鱼(初始体重15 mg)的必需氨基酸需求量。因此开展不同品系,不同生长阶段以及不同饲养条件下的必需氨基酸的平衡模式研究,对于合理构建理想且高效的低蛋白饲料配方具有重要的参考价值。上述大量研究结果是建立在生长需求条件下的,关于必需氨基酸对罗非鱼的免疫系统的营养调控研究相对不足。必需氨基酸在调节营养代谢、免疫力等生理过程中具有多种独特的功能。蛋白质营养不良会降低动物的免疫性能,增加感染疾病的机率(Li等,2007b,Li等,2007a;蔡春芳等,2001)。因此,从免疫系统的营养需求角度进一步深化罗非鱼对必需氨基酸以及其他功能性氨基酸的营养需求研究,构建氨基酸调控罗非鱼免疫功能和抗病力的调控方案,将会在理论指导和生产实践上产生更加积极的作用。
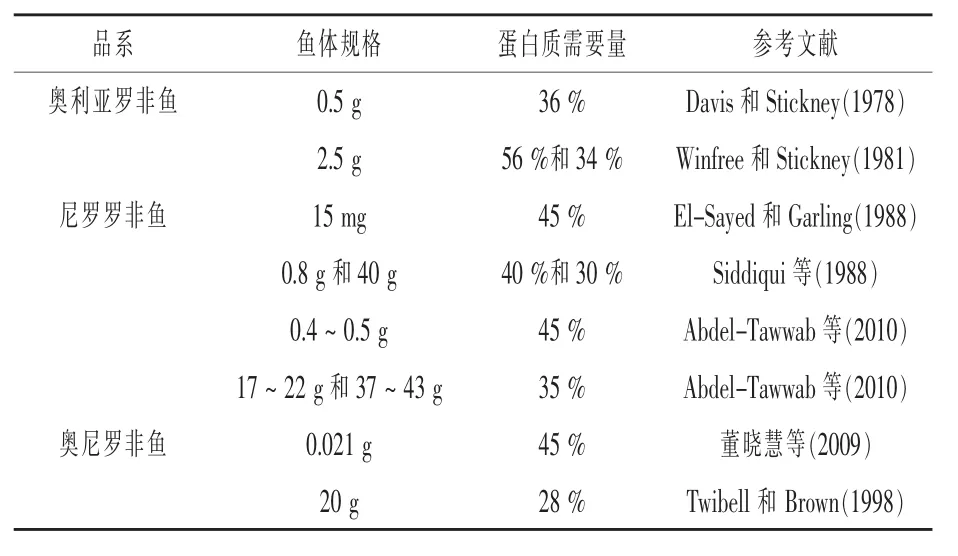
表1 罗非鱼蛋白质需要量
2 脂质以及脂肪酸
脂质包括油脂(甘油三脂)和类脂(磷脂、蜡、萜类、甾类)。饲料中脂质成分是鱼体必需脂肪酸的唯一来源,因此饲料中必须要有适当脂质含量以维持鱼体正常生长、健康和发育。此外,脂质具有多种生理功能,是脂溶性维生素的载体,可以促进其在生物体内的吸收。脂质中的磷脂能够维持细胞结构、膜弹性及渗透性,是类固醇激素和前列腺素的合成前体,同时能够改善饲料风味,影响饲料质地以及鱼体脂肪酸组成等(Lim和Webster,2006)。

表2 尼罗罗非鱼必需氨基酸需要量%
2.1 脂质水平 脂质是鱼类能量的主要来源,但是对于杂食性的罗非鱼而言其需求量相对较低,这主要是由于相对于冷水性鱼类或肉食性鱼类而言,罗非鱼可以更好地利用饲料中的碳水化合物。不同品系罗非鱼对脂质的需求量存在差异。Hanley(1991)研究发现,尼罗罗非鱼对脂质的最低需求量为5.1%。Chou和Shiau(1996)以5种不同脂质含量饲粮(0%、5%、10%、15%和20%,脂质由玉米油、鳕鱼肝油和猪油以1∶1∶1混合组成)投喂奥尼罗非鱼,结果表明,罗非鱼最低脂质需求量为5%,当饲料中脂质水平为12%时达到最佳生长。然而,奥尼罗非鱼对饲料中的脂肪耐受程度是有限的,当饲料中的脂质含量高于12%时,生长发育显著下降,同时机体的脂肪含量显著增加(Jauncey,2000)。此外,脂肪源也会对罗非鱼的脂质需求量产生影响,研究发现,当饲料中含10%鲱鱼油时,奥利亚罗非鱼可以获得最佳生长性能(Stickney 和 Wurts,1986)。
2.2 脂肪酸 研究表明,尼罗罗非鱼、奥利亚罗非鱼和齐氏罗非鱼具有脂肪酸去饱和化以及将18∶2(n-6)和 18∶3(n-3)系列脂肪酸碳链延长为长链n-6和n-3系列多不饱和脂肪酸的能力,当饲料中添加 18∶2(n-6)和 18∶3(n-3)系列脂肪酸时,能够增加鱼体内的长链多不饱和脂肪酸的水平(Visentainer等 2005;Tocher等 2002)。 尼罗罗非鱼脂肪酸的转化能力取决于饲料中的脂肪酸组成。Tocher等(2002)研究认为,给罗非鱼投喂植物油饲料,肝脏脂肪酸去饱和化的活性是投喂鱼油组饲料的两倍,并且长链脂肪酸对脂肪酸去饱和化酶具有抑制作用。相反,当二十碳和二十二碳多不饱和脂肪酸水平低于需求量时,可以诱导脂肪酸去饱和化酶的生成。Kanazawa等(1980)对齐氏罗非鱼研究发现,当饲料中分别添加18∶2(n-6)或者18∶2(n-3)系列脂肪酸时,可增加全鱼极性脂肪酸即 20∶4(n-3)和 22∶6(n-3)脂肪酸的含量。在对尼罗罗非鱼的研究中发现,在全植物蛋白源饲料中,利用亚麻子油[富含 18∶3(n-3)系列脂肪酸]来替代饲料中的葵花籽油[富含 18∶3(n-6)系列脂肪酸],可以显著增加肌肉中的18∶3(n-3)、20∶5(n-3)和 22∶6(n-3)系列脂肪酸的含量(Visentainer等,2005)。 Tocher等(2002)研究发现,尽管投喂植物油饲料,罗非鱼体中 18∶3(n-3)脂肪酸去饱和化含量仍增加,但这种去饱和化的能力非常有限,不能够使组织中 20∶5(n-3)和 22∶6(n-3)脂肪酸的含量与投喂鱼油饲料组含量一致。对齐氏罗非鱼和尼罗罗非鱼的研究中发现两种鱼对n-6系列脂肪酸的需求量分别为1.0%和0.5%(Takeuchi等 1983a;Kanazawa等 1980b)。 在对脂肪源的研究中发现,亚麻酸(n-3)系列脂肪酸[包括 18∶3(n-3),20∶5(n-3),或 22∶6(n-3)系列脂肪酸]和亚麻酸(n-6)系列脂肪酸一样均是尼罗罗非鱼、奥利亚罗非鱼及奥尼罗非鱼的必需脂肪酸(Yildirim-Aksoy 等 2007;El-Sayed 等 2005;Chou等2001)。然而,关于罗非鱼对n-3系列脂肪酸的最佳需要量仍鲜见报道。Yildirim-Aksoy等(2007)研究认为,当饲料中n-6或者n-3系列脂肪酸的含量较高时,可能会对其他脂肪酸产生节约作用。然而,在对齐氏罗非鱼的研究中发现,当饲料中18∶3(n-3)系列脂肪酸水平过高 (大约5%),即严重缺乏n-6系列脂肪酸时会导致鱼体生长速率下降(Kanazawa等,1980b)。同样在对尼罗罗非鱼的研究中也发现,5%鳕鱼肝油[富含20∶5(n-3)和 22∶6(n-3)系列脂肪酸]饲料组的罗非鱼生长较投喂玉米油或黄豆油饲料组慢。Yildirim-Aksoy等(2007)研究报道,投喂鲱鱼油饲料组罗非鱼的红细胞和白细胞总数较高,黏液及抗应激能力均有所提高。投喂缺乏n-6和n-3系列不饱和脂肪酸饲料组的罗非鱼会出现厌食、生长速率下降、饲料效率降低、全鱼的 18∶1(n-9)和 20∶3(n-9)系列脂肪酸含量上升,脂肪肝以及繁殖力下降等症状(NRC 1993)。不同品系的罗非鱼对脂肪酸的需要量见表3。

表3 罗非鱼对脂肪酸的需要量
3 碳水化合物
碳水化合物不仅参与鱼体组织的构成,如五碳糖参与构成细胞核酸,半乳糖是构成神经组织的必需物质,糖蛋白参与形成细胞膜等,而且糖类是鱼类生命活动所需能量的一个重要来源,是饲料中来源广泛且经济的能源,如能合理充分利用,可以部分降低蛋白质作为能量的消耗,起到节约蛋白质的作用,达到降低饲料成本的目的。但鱼类营养学的研究证明,饲料中碳水化合物的含量不宜过高,否则会导致鱼类形成脂肪肝。有研究表明,罗非鱼对不同性质的糖源利用效率存在差异,对淀粉的利用优于葡萄糖 (Shiau和Chen,1993;Shiau 和 Lin,1993)。 Shiau 和 Chuang(1995)研究发现,奥尼罗非鱼对双糖的利用率高于葡萄糖,但低于淀粉,双糖中麦芽糖利用率最佳,其次为蔗糖及乳糖。Lin和Shiau(1995)研究发现,投喂淀粉饲料组罗非鱼,肝脏中碳水化合物代谢相关酶包括苹果酸酶、葡萄糖六磷酸去氢酶及磷酸葡萄糖酸脱氢酶等活性均较投喂葡萄糖饲料组罗非鱼的高,说明罗非鱼肝脏中碳水化合物代谢酶的活性受糖源的性质影响。Wang等(2005)试验表明,在饲料蛋白质含量为29%,脂肪含量为10%时,22%生玉米淀粉就能够满足罗非鱼的能量需要,当碳水化合物水平超过22%时,并不能进一步促进罗非鱼生长,同时研究还发现,罗非鱼对碳水化合物的耐受量较高,当饲料中碳水化合物水平达到46%时,不会限制罗非鱼的生长。
罗非鱼对碳水化合物的利用能力受到多种因素的影响,包括摄食频率、铬以及鱼体规格。在摄食频率方面,Shiau和Lei(1999)通过比较连续投喂及每天投喂两次两种投喂方式对奥尼罗非鱼的碳水化合物利用影响,研究发现,连续喂食可增加奥尼罗非鱼对碳水化合物的利用能力。关于铬对罗非鱼碳水化合物利用的影响,研究结论不尽相同。Shiau和Lin(1993)研究认为,氯化铬可显著改善罗非鱼对葡萄糖的利用率。其后,Shiau和Chen(1993)研究指出,氧化铬比氯化铬更能改善罗非鱼对葡萄糖的利用率。Shiau和Liang(1995)研究报道,饲料中氧化铬能改变罗非鱼对葡萄糖的利用率,并影响饲料中其他营养素的消化率。Shiau和Shy(1998)研究指出,对奥尼罗非鱼而言当葡萄糖饲料中氧化铬含量为204.4 mg/kg时,罗非鱼的生长和对葡萄糖利用能力最强。然而,Pan等(2003)对奥尼罗非鱼的研究认为,饲料中添加2 mg/kg铬并不影响罗非鱼生长及对碳水化合物的利用。此外,不同生长阶段的罗非鱼其对碳水化合物的利用能力也存在差异。Tung和Shiau(1993)比较了两种规格的罗非鱼(4.55 g及0.46 g)对碳水化合物(葡萄糖和淀粉)的利用能力,结果表明,当以葡萄糖为糖源时,大规格罗非鱼的增重率、饲料转换率、蛋白质及能量存积率均优于小规格罗非鱼;然而,当以淀粉为糖源时,各饲料组之间均无明显差异。
罗非鱼对碳水化合物的代谢机制一直是营养研究的热点问题。研究发现,罗非鱼消化系统简单,且缺乏纤维素分解酶。Lin等(1995)研究发现,投喂葡萄糖饲料组的罗非鱼血浆胰岛素浓度高于投喂淀粉饲料组的罗非鱼。罗非鱼摄取不同性质的碳水化合物会影响其氨的排出和氧气的消耗。Shiau和Cheng(1999)研究发现,饲料中添加33%、37%和41%淀粉或葡萄糖,淀粉组罗非鱼的氨排放量较葡萄糖组低,并且无论淀粉组还是葡萄糖组罗非鱼氨排放量随碳水化合物含量的增加而减少;淀粉组总氧消耗量显著高于葡萄糖组;并且,不论淀粉还是葡萄糖,37%和41%碳水化合物组的氧消耗量均高于33%碳水化合物组。
4 维生素
维生素是维持鱼类健康、促进鱼类生长发育和调节生理功能所必需的一类低分子有机化合物,对维持鱼类的代谢过程和生理机能,有着极其重要且不能为其他营养物质所替代的作用。罗非鱼对维生素的需要量虽然很少,但绝大多数维生素不能在体内合成或大量储存在组织中,所以必须由饲料来供给。只有部分维生素能够自身合成。研究发现,罗非鱼胃肠道细菌可产生足够量的维生素B12来满足需求,因此无需在饲料中额外添加维生素 B12(Shiau 和 Lung,1993)。罗非鱼的维生素需要量见表4。

表4 罗非鱼维生素需要量
如果罗非鱼长期维生素摄入不足,不能满足机体对维生素的生理需要,就会导致生理以及代谢障碍,从而影响正常的生理功能,严重时出现维生素缺乏症。罗非鱼的缺乏维生素B2典型症状为厌食、生长迟缓、高死亡率、鳍部溃烂、缺乏正常体色及白内障等。红色奥尼罗非鱼稚鱼缺乏维生素B1时,其生长性能、饲料效率及血细胞比容均会显著下降 (Lim和LesMaster,1991)。 罗非鱼缺乏烟碱酸会产生出血、口鼻部畸形、鳃浮肿及皮肤、鳍与口部溃烂等症状。奥利亚罗非鱼缺乏泛酸会产生生长迟缓、溶血、呆滞、高死亡率、贫血及鳃外围薄层上皮细胞严重增生等症状 (Soliman和Wilson,1992b)。罗非鱼缺乏维生素C会出现骨骼弯曲、腹水、皮肤出血、鳃盖不全、成长减缓、厌食、软骨异常、胶原合成受阻、伤口愈合能力下降、易受疾病感染与高死亡率等症状。在对罗非鱼维生素A的营养需求研究中发现,饲料中添加维生素A不会对罗非鱼免疫系统产生影响(Guo等,2009)。
罗非鱼对维生素的需要量受配方组成的影响。在对奥尼罗非鱼稚鱼研究中发现,饲料中碳水化合物性质会对烟酸的需要量产生影响,当饲料中碳水化合物为葡萄糖时罗非鱼的烟酸最适需要量为26 mg/kg;当为糊精时,其对烟酸的最适需要量为 121 mg/kg(Shiau 和 Suen,1992)。 饲料中蛋白质含量同样会影响罗非鱼维生素B6的需要量。Shiau和Hsieh(1997)研究指出,淡水养殖环境中,当奥尼罗非鱼饲料中蛋白质含量分别为28%和36%时,维生素B6的需要量分别为1.7~9.5 mg/kg和15.0~16.5 mg/kg;养殖于海水中的红色奥尼罗非鱼投喂38%蛋白质饲料时,其维生素B6的最佳需要量为3 mg/kg(Lim等,1995)。Roem等(1990a、b)研究发现,饲料中蛋氨酸含量较高时可满足尼罗罗非鱼对胆碱的需求。甜菜碱可完全取代尼罗罗非鱼对胆碱的需求(Kasper等,2002)。 Soliman 和 Wilson(1992b)研究认为,奥利亚罗非鱼对泛酸的最低需要量为10 mg/kg。然而,Roem等(1990b)研究发现,养殖于循环系统中的罗非鱼,其养殖水体中细菌所提供的泛酸即可满足罗非鱼的需求。同样水体的盐度也会影响罗非鱼对维生素B2的需求量,养殖于淡水中的奥利亚罗非鱼稚鱼及养殖于盐度为32‰c海水中的红色奥尼罗非鱼对维生素B2的需要量分别为6 mg/kg和 5 mg/kg (Lim 等 ,1993;Soliman 和 Wilson,1992a)。当饲料中脂质含量为3%和6%时,奥利亚罗非鱼对维生素E最适需要量分别10 mg/kg和25 mg/kg,即当饲料中添加1%脂质时需添加维生素E3~4 mg(Roem 等,1990c)。 当饲料中添加5%脂质时,尼罗罗非鱼的维生素E需要量为50~100 mg/kg,若脂质增加到10%~15%,则需要量增加至 500 mg/kg(Satoh 等,1987)。 Shiau和Shiau(2001)研究发现,奥尼罗非鱼的维生素E需求量也受饲料中脂肪水平的影响,当饲料脂质含量为5%和12%时,罗非鱼的维生素E需求量分别为42~44 mg/kg和60~66 mg/kg。Huang和Huang(2004)研究指出,喂食氧化脂质(98 meq/kg)的奥尼罗非鱼,维生素E的需求量为62.5 mg/kg,这与Shiau(2001)的研究结果相近。此外,有研究表明,维生素C可节约罗非鱼对维生素E需求量(Shiau 和 Hsu,2002)。
5 矿物质
矿物质是鱼类维持机体渗透压、酸碱平衡等正常代谢活动所必需的营养素。Dato-Cajegas和Yakupitiyage(1996)研究认为,由于罗非鱼可以通过皮肤和鳃盖从外界水域中吸收部分无机盐,在半精养的模式下,一些常量元素(Mg、Na和K)和微量元素 (Zn、Fe、Mn和I) 并不需要在饲料中添加。目前,关于罗非鱼对矿物质的需求量主要集中在钙、磷、镁、锌、钾、铁、铜、钠等(表5)。 在低钙养殖水环境中,奥利亚罗非鱼饲料中含钙7.0 g/kg和磷5.0 g/kg有助于鱼体正常成长及骨头矿化(Robinson等,1987)。对尼罗罗非鱼而言,维持最大生长速率及正常骨骼矿化的磷需求量低于9 g/kg(Watanabe等,1980)。奥利亚罗非鱼饲料中含有500 mg/kg镁及30 mg/kg锌有助于其正常生长;尼罗罗非鱼对镁及锌的需要量分别为590~770 mg/kg及 30 mg/kg(Eid 和 Ghonim,1994;Reigh 等,1991;Dabrowska 等,1989;McClain 和 Gatlin,1988)。 奥尼罗非鱼对钾的最适需要量为2~3 g/kg(Shiau和Hsieh,1997)。 Shiau 和 Ning(2003)研究认为,奥尼罗非鱼铜的需要量为4 mg/kg。罗非鱼铁的需要量受剂型的影响,使用三价铁(柠檬酸铁)及二价铁(硫酸亚铁)时,罗非鱼对铁的需要量分别为150~160 mg/kg和85 mg/kg,另外,有研究报道,罗非鱼体内柠檬酸铁的生物效价为硫酸亚铁的49%(Shiau和Su,2003)。淡水养殖奥尼罗非鱼钠的需要量为1.6 g/kg,海水养殖罗非鱼不必在饲料中额外添加钠(Shiau 和 Lu,2004)。
6 总结与展望
罗非鱼作为近年来国内产量增长速度较快的品种之一,具有较好的发展前景。目前在我国养殖的罗非鱼品系相对较多,如尼罗罗非鱼、奥利亚罗非鱼、奥尼罗非鱼以及吉富罗非鱼,由于罗非鱼品系之间在营养素的需求上存在较大差异,所以对单一品系以及品系之间营养需求差异进行系统的研究,对于指导开发不同品种的罗非鱼配合饲料,促进养殖产业的健康、高效发展具有重要的意义。另外,随着罗非鱼规模化、集约化养殖业的发展,罗非鱼的肉质愈来愈受到人们的重视。因此,罗非鱼营养需求的研究不只局限于营养水平对鱼类生长和生理的影响,应该利用分子生物学技术,深入研究蛋白质(氨基酸)、脂肪(脂肪酸)、碳水化合物等营养物质的营养生理代谢规律,阐明鱼肉品质的形成机理并实现营养调控,从而使罗非鱼肉质以及风味得到维护和改善,这将有利于罗非鱼产业的健康发展。
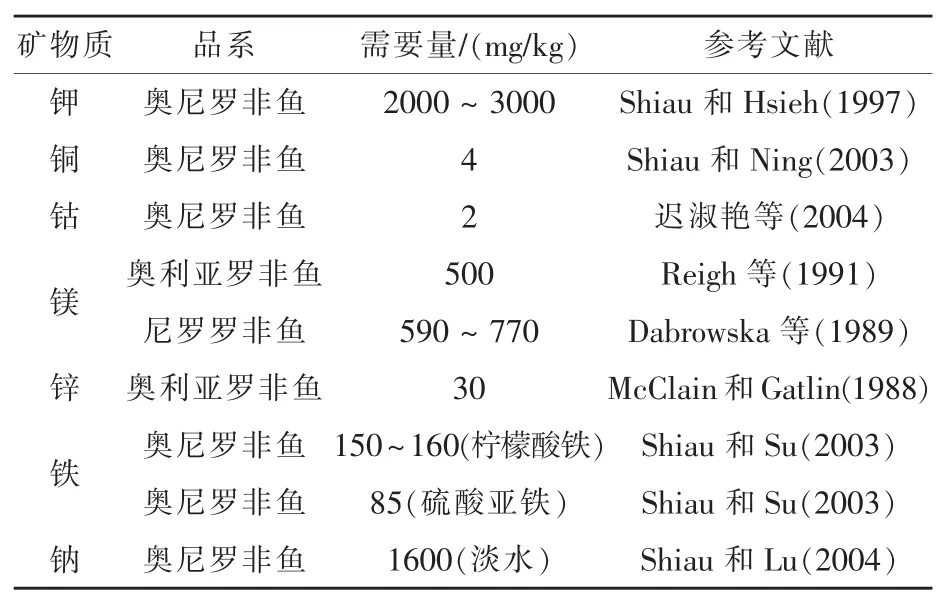
表5 罗非鱼对矿物质需要量
[1]蔡春芳,吴康,潘新法,等.蛋白质营养对异育银鲫生长和免疫力的影响[J].水生生物学报.2001,25(6):590~595.
[2]迟淑艳,周歧存,杨奇慧,等.罗非鱼营养研究进展[J].饲料研究,2004,9:10~14.
[3]董晓慧,耿旭,郭云学,等.奥尼罗非鱼仔稚鱼饲料中适宜蛋白质水平的研究[J].中国饲料,2009,7:29~32
[4]Abdel-Tawwab M,Ahmad M H,Khattab Y A E,et al.Effect of dietary protein level,initial body weight,and their interaction on the growth,feed utilization,and physiological alterations of Nile tilapia,Oreochromis niloticus(L)[J].Aquaculture,2010,298:267~74.
[5]Chou B S,Shiau S Y.Optimal dietary lipid level for growth of juvenile hybrid tilapia,Oreochromis niloticus × Oreochromis aureus[J].Aquaculture,1996,143:185~195.
[6]Chou B S.Shiau S Y.Both n-6 and n-3 fatty acids are required for maximal growth of juvenile hybrid tilapia[J].North American Journal of Aquaculture,1999,61:13~20.
[7]Dato-Cajegas C R S,Yakupitiyage A.The need for dietary mineral supplementation for Nile tilapia,Oreochromis niloticus,cultured in a semiintensive system[J].Aquaculture,1996,144:227~237.
[8]Eid A E,Ghonim S I.Dietary zinc requirement of fingerling Oreochromisniloticus[J].Aquaculture,1994,119:259~264.
[9]El-Sayed A F M,Garling Jr D L.Carbohydrate-to-lipid ratios in diets for tilapia fingerlings[J].Aquaculture,1988,73:157~163.
[10]El-Sayed A F M,Mansour C R,Ezzat A A.Effects of dietary lipid source on spawning performance of Nile tilapia (Oreochromisniloticus)broodstock reared at different water salinities[J].Aquaculture,2005,248:187~196.
[11]Hu C J,Chen S M,Pan C H.et al.Effects of dietary vitamin A or bcarotene concentrations on growth of juvenile tilapia,Oreochromis niloticus×Orechromis aureus[J].Aquaculture,2006,253:602~607.
[12]Huang C H,Huang S L.Effect of dietary vitamin E on growth,tissue lipid peroxidation,and liver glutathione level of juvenile hybrid tilapia,Oreochromis niloticus× O.aureus,fed oxidized oil[J].Aquaculture,2004,237:381~389.
[13]Kasper C S,White M R,Brown P B.Betaine can replace choline in diets for juvenile Nile tilapia,Oreochromis niloticus[J].Aquaculture,2002,205:119~126.
[14]Li P,Gatlin D M,Neill W H.Dietary supplementation of a purified nucleotide mixture transiently enhanced growth and feed utilization of juvenile red drum,Sciaenops ocellatus[J].Journal of the World Aquaculture Society,2007b,38:281~286.
[15]Li P,Yin Y L,Li D,et al.Amino acids and immune function[J].British Journal of Nutrition,2007a,98(2):237~252.
[16]Lim C,Webster C D.Nutrient requirements[M].Tilapia:biology,culture and nutrition.Haworth Press,Binghamton,New York,2006.469~501.
[17]Lin J H,Shiau S Y.Hepatic enzyme adaptation to different dietary carbohydrates in juvenile tilapia (Oreochromis niloticus × O.aureus)[J].Fish Physiology and Biochemistry,1995,14:165~170.
[18]Pan Q,Liu S,Tan Y,et al.The effect of chromium picolinate on growth and carbohydrate utilization in tilapia Oreochromis niloticus×O.aureus[J].Aquaculture,2003,225:421~429.
[19]Robinson E H,Rawles S D,Yette H E,et al.An estimate of the dietary calcium requirement of fingerling Tilapia aureus reared in calcium-free water[J].Aquaculture,1984,41:389~393.
[20]Shiau S Y,Cheng D J.Ammonia excretion and oxygen consumption of tilapia are affected by different carbohydrate ingestion[J].Fisheries Science,1999,65:321~322.
[21]Shiau S Y,Chin Y H.Estimation of the dietary biotin requirement of juvenile hybrid tilapia,Oreochromis niloticus × O.aureus[J].Aquaculture,1999,170:71~78.
[22]Shiau S Y,Chuang J C.Utilization of disaccharides by juvenile tilapia,Oreochromis niloticus × O.aureus[J].Aquaculture,1995,133,249~256.
[23]Shiau S Y,Hsieh H L.Vitamin B6requirements of tilapia Oreochromis niloticus×O.aureus fed two dietary protein concentrations[J].Fisheries Science,1997,63:1002~1007.
[24]Shiau S Y,Hsu C Y.Vitamin E sparing effect by dietary vitamin C in juvenile hybrid tilapia,reochromis niloticus × O.aureus[J].Aquaculture,2002,210:335~343.
[25]Shiau S Y,Hsu T S.Quantification of vitamin C requirement for juvenile hybrid tilapia,Oreochromis niloticus × O.aureus,with L-ascorbyl-2-monophosphate-Na and L-ascorbyl-2-monophosphate-Mg[J].Aquaculture,1999,175:317~326.
[26]Shiau S Y,Huang S Y.Dietary folic acid requirement for maximal growth of juvenile tilapia,Oreochromis niloticus × O.aureus[J].Fisheries Science,2001,67:655~659.
[27]Shiau S Y,Hwang J Y.Vitamin D requirement of juvenile hybrid tilapia Oreochromis niloticus× O aureus[J].Nippon Suisan Gakkaishi,1993,59:553~558.
[28]Shiau S Y,Jan F L.Dietary ascorbic acid requirement of juvenile tilapia Oreochromis niloticus × O.aureus [J].Nippon Suisan Gakkaishi,1992,58:671~675.
[29]Shiau S Y,Liang H S.Carbohydrate utilization and digestibility by tilapia,Oreochromis niloticus × O.aureus,are affected by chromic oxide inclusion in the diet[J].The Journal of Nutrition,1995,125:976~982.
[30]Shiau S Y,Lin S F.Effect of supplemental dietary chromium and vanadium on the utilization of different carbohydrates in tilapia,Oreochromis niloticus × O.aureus[J].Aquaculture,1993,110:321~330.
[31]Shiau S Y,Lo P S.Dietary choline requirements of juvenile hybrid tilapia,Oreochromis niloticus × O.aureus[J].The Journal of Nutrition,2000,130:100~103.
[32]Shiau S Y,Ning Y C.Estimating of dietary copper requirements for juvenile tilapia,Oreochromis niloticus× O.aureus[J].Animal Science,2003,77:287~292.
[33]Shiau S Y,Shiau L F.Reevaluated of the vitamin E requirement of juvenile tilapia,Oreochromis niloticus× O.aureus[J].Animal Science,2001,72:529~534.
[34]Shiau S Y,Shy S M.Dietary chromic oxide inclusion level required to maximize glucose utilization in hybrid tilapia,Oreochromis niloticus × O.aureus[J].Aquaculture,1998,161:357~364.
[35]Shiau S Y,Su L W.Ferric citrate is half as effective as ferrous sulfate in meeting the iron requirement of juvenile tilapia,Oreochromis niloticus × O.aureus[J].The Journal of Nutrition,2003,133:483~488.
[36]Shiau S Y,Su S L.Juvenile tilapia(Oreochromis niloticus× O.aureus)requires dietary myo-inositol for maximal growth[J].Aquaculture,2005,243:273~277.
[37]Shiau S Y,Suen,G S.Estimation of the niacin requirements for tilapia fed diets containing glucose or dextrin[J].The Journal of Nutrition,1992,122:2030~2036.
[38]Shiau SY,Lei M S.Feeding strategy does affect carbohydrate utilization by hybrid tilapia Oreochromis niloticus × O.aureus[J].Fisheries Science,1999,65:553~557.
[39]Shiau S Y,Su S L.Juvenile tilapia (Oreochromis niloticus× Oreochromis aureus)requires dietary myo-inositol for maximal growth[J].Aquaculture,2005,243(1~4):273~277.
[40]Shiau S Y,Hsu T S.L-Ascorbyl-2-sulfate has equal antiscorbutic activity as L-ascorbyl-2-monophosphate for tilapia,Oreochromis niloticus × O.aureus[J].Aquaculture,1995,133:147~157.
[41]Soliman A K,Jauncey K,Roberts R J.Water-soluble vitamin requirements of tilapia:ascorbic acid (vitamin C)requirement of Nile tilapia,Oreochromis niloticus(L)[J].Aquaculture and Fisheries Management,1994,25:269~278.
[42]Tocher D R,Angaba M,Hastings N,et al.Nutritional regulation of hepatic fatty acid desaturation and polyunsaturated fatty acid composition in zebrafish(Danio rerio)and tilapia (Oreochromis niloticus)[J].Fish Physiology and Biochemistry,2002,24:309~320.
[43]Wang Y,Liu Y J,Tian L X.Effects of dietary carbohydrate level on growth and body composition of juvenile tilapia,Oreochromis niloticus×O.aureus[J].Aquaculture Research,2005,36(14):1408~1413.
[44]Winfree R A,Stickney R R.Effects of dietary protein and energy on growth,feed conversion efficiency and body composition of Tilapia aureus[J].The Journal of Nutrition,1981,111:1001~1012.
[45]Yildirim-Aksoy M,Lim C,Davis D A,et al.Influence of dietary lipid sources on the growth performance,immune response and resistance of Nile tilapia (Oreochromis niloticus)to Streptococcus iniae challenge[J].Journal of Applied Aquaculture,2007,19:29~49.
