长筒石蒜生物量构成和养分质量分数季节动态1)
2012-07-02鲍淳松张鹏翀张海珍黎念林
鲍淳松 张鹏翀 张海珍 江 燕 黎念林
(杭州植物园,杭州,310013)
长筒石蒜(Lycoris longituba)隶属石蒜科(Amaryllidaceae)石蒜属(Lycoris Herb.),为具地下鳞茎的多年生草本植物,春季前就有部分出叶[1],花白色,花期7—8月份,果期8—11月份,花叶期分离。石蒜属植物是一种极具开发潜力的球根花卉,为了满足日益扩大的市场需求,近年来,部分研究者对石蒜属植物进行了一些施肥方面的研究[2-3]。笔者[1,4-6]发现对石蒜属植物施肥未显著增加鳞茎产量。为了探究其原因,有必要摸清石蒜属植物的养分质量分数、分配及其季节动态,但至今对石蒜属植物矿物质养分质量分数、碳转化、转移和分布的动态机理研究尚很有限。为了探索石蒜类植物的养分机理,本研究以长筒石蒜为研究对象,对其生物量构成和地上、地下部分N、P、K质量分数进行动态测定,为长筒石蒜的生态学机理研究和科学施肥提供理论依据,同时为石蒜属植物的研究和应用提供基础数据。
1 试验地概况
材料来自杭州植物园种质资源圃(120°16'E,30°15'N),圃地为红壤,土壤全氮质量分数 0.2 g·kg-1,全磷质量分数 0.26 g·kg-1,全钾质量分数 4.1 g·kg-1,有机质质量分数 15 g·kg-1,pH 值 5.6。年平均气温16.1℃,极端最低气温-10.5℃,极端最高气温41℃,年平均降水量1 400 mm。
2 材料与方法
供试物种为长筒石蒜,该批鳞茎种植于2005年,种植时的株行距为10 cm×15 cm,畦宽90 cm。
植株取样采用整株挖掘法。测定时间从2009年4月份至2011年4月份,根据不同的生长期,选择天气晴好的日子,每隔1个月左右挖掘整株植物1次,共计挖掘26次,每次取样随机挖掘2行长筒石蒜,重复3次,作为该群体生长状态的样本,其中2009年10月份,随机挖掘开花结实的大鳞茎9只。挖掘时整株掘起,并尽量保证植株根系完整,随后用自来水冲洗干净,分根系、鳞茎和地上部分3个组分,85℃烘干至恒质量,小型植物样品粉碎机粉碎至粉未,供养分分析用。全N采用凯氏定氮法,全P和全K测定,先采用硫酸—高氯酸消煮法制备待测液,再分别用钼锑抗比色法和原子吸收法测定。
养分再吸收效率(nutrient resorption efficiency,NR,E)按如下公式计算[7-9]:
NR,E=100%×(绿叶养分质量分数—衰老叶养分质量分数)/绿叶养分质量分数。
试验测定数据输入计算机建立数据库,经计算汇总后用EXCEL 2003进行常规分析。
3 结果与分析
3.1 生物量百分比构成季节动态
图1为地上部分、鳞茎和根系的百分比构成动态。根系生物量占比变化呈单底格式,范围为3.1% ~8.6%,出叶前(12月份)根生物量百分比最大,随着叶片生长,根系占比减小,枯叶期(4月份)根系占比达最小值(3.1%),5—6月份平均占 3.7%,与鲍淳松[1]根系占比3.3%的结果基本相符。

图1 长筒石蒜生物量构成百分比季节动态
长筒石蒜叶生物量占比呈单峰格式,在3月份达到最大,占整个生物量的29.6%。开花结实植株在开花结实期10月份地上部分生物量最大,占37.0%,但群体平均占10%左右,即该群体开花鳞茎约占1/4。
鳞茎生物量占比动态则呈双底格式,最小值分别在3月份和10月份,即叶期最低值65.6%,果期最低值57.9%(开花结实植株);最大值在6月份,达 96.3%。
3.2 N、P、K 质量分数变化动态
3.2.1 根系
图2为根系的N、P、K质量分数变化动态。N的质量分数变化比较有规律,即根系生长结束的冬季叶刚出土时(12月份—翌年2月份)质量分数最高,枯叶期(4月份)和开花期(10月份)是2个低谷,期间开花前达到一个小高峰;P的质量分数较低,波动不大,8月份最高,6、9月份较低;而K的质量分数变化不规则,但果期质量分数最高,3月份最低。N 质量分数最高达 41.8 g·kg-1,最低 15.3 g·kg-1,平均27.6 g·kg-1;P 的质量分数较低,最高 8.6 g·kg-1,最低 3.1 g·kg-1,平均 5.1 g·kg-1;K 的质量分数最高 27.2 g·kg-1,最低 12.9 g·kg-1,平均18.5 g·kg-1。年平均质量分数 w(N)∶w(P)∶w(K)=5 ∶1 ∶4。

图2 根系中N、P、K质量分数变化动态
3.2.2 鳞茎
图3为鳞茎中的N、P、K质量分数变化动态。N呈双底形态,N以出叶前(11月份)最高,出叶后一直下降,4月份最低,枯叶后增加,开花结实期间减少后再增加;K的变化格式与N有点类似,4、9月份较低,5、11月份较高;P的质量分数低,叶期呈下降趋势,其余呈波动状,但12月份最高。N质量分数最高 26.3 g·kg-1,最低 7.4 g·kg-1,平均 15.6 g·kg-1;P 的质量分数低,最高5.4 g·kg-1,最低2.1 g·kg-1,平均 3.8 g·kg-1;K 的质量分数最高 12.3 g·kg-1,最低 4.9 g·kg-1,平均 8.9 g·kg-1。年平均质量分数w(N)∶w(P)∶w(K)=4∶1∶2。

图3 鳞茎中N、P、K质量分数变化动态
3.2.3 地上部分
图4为地上部分中的N、P、K质量分数变化动态,其中12月份—翌年4月份为叶的养分质量分数,8—11月份为花果的养分质量分数。出叶时N质量分数最高,达48.5 g·kg-1,随着叶片生长 N 质量分数呈抛物线形下降,4月份达最低,为14.4 g·kg-1,平均 35.8 g·kg-1;地上部分花果 N 质量分数变化不明显,抽苔后到开花N质量分数先增后减,再增加,可能与花葶、果实各器官占比变化有关,平均13.4 g·kg-1。P 的质量分数变化不大,出叶早期和枯叶期及果初期较低,约3.8 g·kg-1,盛叶期前和果成熟期较高,分别为 7.4、8.3 g·kg-1,叶期平均 5.0 g·kg-1,果期平均 5.4 g·kg-1。K 质量分数叶前期略高,盛叶期略低,平均 14.0 g·kg-1;花果期略高,成熟期略低,平均 15.0 g·kg-1。全年 N、P、K 质量分数平均值分别为 25.8、5.1、14.5 g·kg-1,年平均质量分数 w(N)∶w(P)∶w(K)=5 ∶1∶3。

图4 地上部分中N、P、K质量分数变化动态
由于长筒石蒜绿叶期很短,叶枯萎很快,且每年的物候期会因天气等因素有几天甚至更长时间的波动,所以同一个月内养分质量分数会有变化。
3.3 N、P、K 养分总量分配动态
养分总量等于养分质量分数与生物量的乘积,分配指养分在各组成部分之间的构成比例。根和鳞茎中的N、P、K养分总量呈双底格式,地上部分呈单峰格式。
3.3.1 N 总量分配动态
从图5中可看出,鳞茎中N总量在盛叶期最低,占44.2%,花果期次低,占63.4%,叶枯后的5月份最高,占95.7%,全年平均占79.5%。地上部分盛叶期(3月份)N总量占比最大,为47.5%,比鳞茎中的总量还高,最低12月份,占0.7%;花果期最高占28.4%,年平均15.8%。根系中的N总量变化不大,最高12月份,占15.7%,新根生长前后(5月份)较低,为 4.3%,年平均 8.7%。
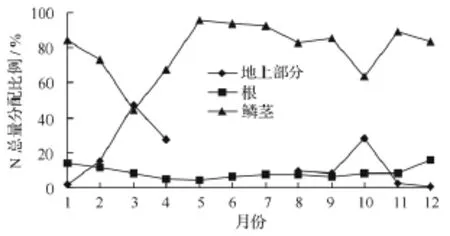
图5 N总量分配动态
3.3.2 P 总量分配动态
从图6中可看出,鳞茎中P总量在叶期最低,占58.0%,叶枯后的5月份最高,占95.3%,全年平均占81.5%。地上部分P总量占比最大在3月份和10月份,各占36.6%,最低在12月份,占0.5%,年平均15.4%。根系中的P总量叶期4月份最低,占3.4%,果熟期10月份最低,占5.3%,最高在出叶早期,占10.7%,年平均占7.0%。

图6 P总量分配动态
3.3.3 K 总量分配动态
从图7看出,鳞茎中K总量在果期最低,占4 4.7% ,盛叶期次低,占49.3%,叶枯后最高,占94.6%,全年平均74.9%。地上部分花果期最大,为46.2%,盛叶期次之,占40.2%,出叶早期12月份最低,占0.7%,年平均19.1%。根系中的K总量以出叶前最高,枯叶期后最低,分别为16.5%和5.4%,年平均10.8%。

图7 K总量分配动态
3.4 养分再吸收
表1是2011年3月8日盛叶期和2011年4月19日枯叶期叶片中N、P、K质量分数。N、P、K再吸收率分别为:46.7%、49.6%、63.4%。

表1 叶片衰老前后的N、P、K质量分数
4 结论与讨论
生物量分配动态:根系生物量占比动态呈单底形态,范围为3.1% ~8.6%,刚出叶时最高,出叶后不断降低,枯叶期达最低,枯叶后,根系占比不断上升,特别是秋季上升最快。经推论:长筒石蒜根系以夏秋季节生长为主,叶期不长根,根叶生长不同期。这种根系生长特性与 Burton 等[10]、赵爱芬等[11]的研究结果有别,与人们对植物常规的认识有别。换言之,绿叶期正是根系的衰老期。Chapin[12]认为,一旦分生组织死亡或休眠,植物就不对提高的养分有效性作反应;根吸收能力随着根龄的增加而下降。根系只有不断生长,才能伸展到新的土壤空间中去吸收更多的有效养分。根据根系生长与养分吸收的关系,在叶期进行施肥正面效果不显著[1,5-6],有可能也与施肥季节有关,即与根系生物学特性有关。
鳞茎生物量占比呈双底形态,最低占57.9%,最高为96.3%,叶生物量占比最高为29.6%。花果生物量最高占37.0%。对开花结实的植株而言,生殖生长消耗的生物量大于叶生物量,外观上,结实后鳞茎变的松软,也说明鳞茎中大量的碳水化合物被转移、消耗。由于鳞茎生于地下,有分球的情况发生,很难确定某个鳞茎能否连年开花,经推论:由于开花结实消耗大量营养,对于长筒石蒜单个鳞茎来说,不能连年开花;而以促成栽培为目的的,应控制开花(摘蕾)以利增加鳞茎产量。
养分质量分数和矿物量分配:根系、鳞茎、叶、花果中N、P、K年平均质量分数分别为w(N)∶w(P)∶w(K)=5 ∶1 ∶4、w(N)∶w(P)∶w(K)=4 ∶1 ∶2、w(N)∶w(P)∶w(K)=7 ∶1 ∶3、w(N)∶w(P)∶w(K)=5∶2∶6。开花结实N、P、K消耗占各自总矿物量的28.4%、36.6%、46.2%,即开花结实 K 和 P损失比N的损失大。盛叶期叶的N、P、K量占各自总矿物量的比率分别为 47.5%、36.6%、40.2%。长筒石蒜生物量、养分质量分数及矿物量的年内变化规律,对于栽培管理实践应有指导意义。
再吸收及生态机制:养分再吸收或再分配是指植物在叶片凋落前把部分营养物质转移到其他器官以再利用,是主要的养分保护机制,不仅提高了植物生长对贫瘠土壤的适应性,还提高了植物在贫瘠土壤中的竞争能力[13-14],影响到竞争、养分吸收和生产等诸过程[15]。
石蒜类植物对养分的再吸收能力较强,从实际观察中发现,石蒜类植物对土壤养分的要求不高,能在相对贫瘠的土壤上生长,就长筒石蒜而言,进入枯叶期,大部分叶片是从叶尖端逐步向下枯萎的,叶枯萎部分极大地收缩、皱曲,而且枯叶极易腐烂分解,这种枯萎方式可以保证更大的养分再转移,枯叶的快速分解,有利于枯叶中存留的养分快速归还土壤。
叶片 N、P、K 再吸收率分别达 46.7%、49.6%、63.4%,推测这是一个保守值,一方面由于前述的枯叶方式,而且叶片成熟过程中养分也可能回收,另外,是因为叶生长期短,质量分数变化快,特别是N,其质量分数下降很快,因此,实际再吸收率可能会更高,有待做更详细的研究。
长筒石蒜根系为须根,二级细根比例很小,本研究未做区分。根据图1中4—12月份根生物量占比的变动幅度(3.1% ~8.6%),大部分根得到了更新,粗略估计,根寿命在2 a以内,根的周转率为0.64次·a-1[16-17]。长筒石蒜根生物量所占的比例小,但根系在绿叶期N、P、K再吸收率分别达到63.4%、47.3%、36.2%。据报道,目前细根的养分内循环研究结论还不一致,有的研究认为树木的细根存在或者可能存在养分内循环,也有研究认为树木的细根基本上不存在这种养分的转移,研究方法是通过比较活细根和死细根中各养分的浓度判定衰老细根是否存在养分转移[18],但死细根中有可能会有微生物活动,有可能发生部分分解,土壤中的养分也极有可能渗入死细胞中,而对长筒石蒜活根的动态研究表明,根的N、P、K再吸收是明显的。
石蒜属植物鳞茎生长缓慢,对土壤要求不高,它们具有缓生种的养分策略[12]。可以肯定,石蒜属植物都应有非常发达的内循环机制,其中鳞茎就是一个养分、碳和水的存贮库,当叶片开始生长时,由“库”提供养分和碳“源”用于构建叶片,同时,叶片开始光合作用,逐渐变为碳源,多余的养分再吸收回鳞茎中,包括叶片开始衰老后大部分养分也回流到鳞茎中,在这段绿叶期,根系中的养分也被部分回收,但鳞茎中的N、P、K因供给而质量分数减少;枯叶后的5月份,根系更新生长,鳞茎N、K质量分数增加,同时花芽开始分化发育[19]。开花结实季节,鳞茎中的养分和碳水化合物又再次作为“源”,用于构建花器、果实、种子,由于花器较大,生长快,鳞茎和根系养分质量分数再次减少。开花结实过程中,花葶、果皮部分为绿色能进行光合作用,种子成熟过程中,营养向种子转移,包括地上绿色部分中的养分也可能直接被未成熟种子吸收利用。种子老熟后,绿色部分向下端逐渐枯萎,可以推断也进行有效的养分和碳水化合物的回收。果实成熟后(秋季),根快速生长,鳞茎中的N、K,根系中的N质量分数也快速增加,但根系中的K质量分数先增后减。出叶后又进入下一个循环。
石蒜属植物叶片蜡质层丰富,叶面施肥无效[4],但这种疏水结构,使雨水不易停留于叶表,从而有效地减少因淋溶而丢失养分。枯叶容易分解,有利于在根系生长期归还土壤供植物吸收。
鳞茎既是养分和非结构性碳的存贮库,也是水分的存贮库、备用库。当受干旱胁迫时,鳞茎先通过牺牲最外层鳞片以保存水分、转移水分、转移营养的方式来延续生命,因此,它很耐干旱,鳞茎越大,在干旱条件下存活时间越长,大的鳞茎几年不浇水,还能恢复生长,但相对来说不耐水湿,短暂的水淹问题不大,但长时间缺氧就易烂根或导致鳞茎腐烂。长筒石蒜的这种生态机制(包括养分机制、碳机制和水分机制),通过鳞茎的中转,使长筒石蒜(石蒜属植物)能够具有诸如养分回收率高、耐瘠薄、耐干旱以及花叶不相见、根叶生长不同步等许多与众不同的生理生态学和生物学特性,值得进一步深入研究。
[1]鲍淳松,朱春艳,张海珍,等.施肥对长筒石蒜生长的效应研究[J].浙江农业科学,2009,(6):1092-1094.
[2]杨志玲,谭梓峰,杨旭,等.施肥对红花石蒜物质积累和分配的影响[J].中南林学院学报,2006,26(6):150-154.
[3]刘志高,黄华宏,吴家胜,等.石蒜鳞茎栽培中施用氮磷钾肥的效应[J].南京林业大学学报:自然科学版,2009,33(2):137-140.
[4]鲍淳松,江燕,冯有林,等.叶面施肥对红蓝石蒜生长的影响[J].江苏农业科学,2008(增刊):135-136,193.
[5]鲍淳松,江燕,冯有林,等.施肥对红蓝石蒜生长的响应初报[J].江西农业大学学报,2009,31(增刊):27-31.
[6]鲍淳松,张海珍,江燕,等.换锦花生长特性及其高施肥量对生长的影响[J].青岛农业大学学报:自然科学版,2010,27(增刊):31-33,37.
[7]李志安,王柏荪,林永标,等.植物营养转移研究进展[J].武汉植物学研究,2000,18(3):229-236.
[8]何维明,张新时.沙地柏对毛乌素沙地3种生境中养分资源的反应[J].林业科学,2002,38(5):1-6.
[9]叶功富,张立华,林益明,等.福建东山短枝木麻黄小枝氮磷含量及其再吸收率季节动态[J].生态学报,2009,29(12):6519-6526.
[10]Burton A J,Pregitzer K S,Hendrick R L.Relationships between fine root dynamics and nitrogen availability in Michigan northern hardwood forests[J].Oecologia,2000,125:389-399.
[11]赵爱芬,黄学文.春小麦根系生长动态研究[J].华北农学报,1993,8(1):88-93.
[12]Chapin F S.The Mineral nutrition of wild plants[J].Annual Review of Ecology and Systematics,1980,11:233-260.
[13]Aerts R.Nutrient resorption from senescing leaves of perennials:are there general patterns?[J].Journal of Ecology,1996,84:597-608.
[14]孙书存,陈灵芝.东灵山地区辽东栎叶养分的季节动态与回收效率[J].植物生态学报,2001,25(1):76-82.
[15]Killingbeck K T.Nutrients in senesced leaves:keys to the search for potential resorption and resorption proficiency[J].Ecology,1996,77(6):1716-1727.
[16]单建平,陶大立.国外对树木细根的研究动态[J].生态学杂志,1992,11(4):46-49.
[17]张小全,吴可红,Dieter Murach.树木细根生产与周转研究方法评述[J].生态学报,2000,20(5):875-883.
[18]黄石竹,张彦东,王政权.树木细根养分内循环[J].生态学杂志,2006,25(11):1395-1399.
[19]李爱荣,周坚.中国石蒜叶片的生长周期及其发育过程的研究[J].植物学通报,2005,22(6):680-686.
