寄主龄期对前裂长管茧蜂寄生与生长发育的影响
2012-06-19陈科伟刘春燕
陈科伟,邵 屯,刘春燕,曾 玲
(1华南农业大学资源环境学院,广东广州510642;2广东清远出入境检验检疫局,广东清远511515;3广东省森林病虫害防治与检疫总站,广东广州510173)
前裂长管茧蜂Diachasmimorpha longicaudata(Ashmead)属膜翅目Hymenoptera茧蜂科Bracoindae,主要分布于印度-环太平洋地区,是许多实蝇类害虫,如橘小实蝇Bactrocera dorsalis(Hendel)、地中海实蝇Ceratitis capitata(Wiedemann)、加勒比海实蝇Anastrephpa suspense(Loew)等幼虫—蛹期的重要寄生性天敌[1-3].由于该蜂在室内易于饲养繁殖,自20世纪30年代以来,已陆续被世界范围内许多国家和地区交互引进[4-7].华南农业大学资源环境学院昆虫学系于2004年8月从泰国引进前裂长管茧蜂,对该蜂的生物学、生态学特性及繁殖技术进行了研究[8-9],实现了一定规模的批量繁殖,并进行了初步的田间释放试验[10].目前,有关橘小实蝇幼虫龄期对前裂长管茧蜂寄生与生长发育的影响鲜见系统的研究报道.深入研究寄生蜂对寄主龄期的选择与寄主适合性的关系,有助于认识寄生蜂与寄主之间的相互作用,并为定量评估寄生蜂的控害潜能及大量繁殖提供相关的科学依据.
1 材料与方法
1.1 供试蜂种与虫源
橘小实蝇Bactrocera dorsalis采自广州市天河区杨桃公园,通过采集受害杨桃获得,带回室内饲养,将老熟幼虫放置于细沙中令其化蛹,细沙含水量(w)保持10% ~15%,7~10 d后沙蛹分离,蛹移入50 cm×50 cm×60 cm的养虫笼中待其羽化,收集橘小实蝇成虫.
橘小实蝇在室内主要用人工饲料进行繁育,幼虫人工饲料配方(由泰国农业部生物防治研究所提供):香蕉300 g,新鲜玉米300 g,酵母30 g,蔗糖30 g,苯甲酸钠 0.6 g,纤维 30 g,盐酸 1.2 mL,蒸馏水300 mL.
前裂长管茧蜂 Diachasmimorpha longicaudata,2004年7月从泰国农业部生物防治研究所引进,在室内用橘小实蝇作为寄主进行繁育.
1.2 试验条件
养虫室温度为(26±1)℃,相对湿度为60% ~75%,光照周期为14 h光:10 h暗.
1.3 试验设备
人工气候箱型号为PYX-300Q-B(广东韶关科力试验仪器有限公司),误差为±1℃.
养虫笼:①笼子规格为50 cm×50 cm×60 cm,框架、底座和前面为不锈钢材料,其余面为铁纱网;顶部前倾45°,封有透明玻璃;前面有2个直径为10 cm的圆形开口,带有80 cm长、100目的纱网袖套,以便更换清水和饲料;养虫笼侧面为拉门,以便清理.②自制的小型养虫笼,底面直径为15 cm,高为30 cm,由塑料围成密闭的圆柱体,上表面接80目沙网袖口.
1.4 试验设计
选取羽化后3 d并已交配的雌蜂80头,分成3组,每组20头,分别提供1龄幼虫(2日龄)、2龄幼虫(4日龄)、3龄幼虫初(5~6日龄)和3龄末(7日龄)的橘小实蝇幼虫200头,6 h后将幼虫移出,按正常繁蜂程序进行饲养.每24 h解剖1次,每次解剖30头被寄生幼虫,观察记录各处理前裂长管茧蜂各虫态的存活率及发育历期,待羽化后统计雌雄比例.每处理设5个重复.
前裂长管茧蜂个体大小的测量:衡量寄生蜂个体大小的常用指标有头宽、体长、腹宽及后足胫节长度等,其中后足胫节骨化程度高,不易扭曲、变形,其长度与个体大小呈正相关.本试验选用后足胫节长度(Hind tibia length)来代表个体大小,每种处理分别测量30头以上雌、雄个体的后足胫节长度.

2 结果与分析
2.1 前裂长管茧蜂对不同龄期橘小实蝇幼虫的寄生效能
寄主龄期对前裂长管茧蜂的寄生效能有明显的影响(图1).前裂长管茧蜂对3龄初橘小实蝇幼虫的寄生行为积极主动,在试验观察的6 h内有85.50头幼虫被寄生.其次为2龄幼虫,6 h内有63.00头被寄生.寄主龄期过低或过高对前裂长管茧蜂的寄生行为均有一定的抑制作用.6 h内1龄末与3龄末橘小实蝇幼虫仅有35.60与33.21头被寄生.

图1 前裂长管茧蜂6 h内对不同龄期橘小实蝇幼虫的寄生数量Fig.1 The number of Bctrocera dorsalis larvae parasited by Diachasmimorpha longicaudata within 6 hours
2.2 寄主年龄对前裂长管茧蜂生长发育的影响
寄主龄期对前裂长管茧蜂的发育速率有明显的影响,前裂长管茧蜂各虫态的发育速率均随寄主龄期的增加而逐步减缓(表1).其中,对卵发育的延缓作用最为明显.以1龄橘小实蝇幼虫为寄主时,前裂长管茧蜂卵的历期为1.8 d,以2龄、3龄初及3龄末橘小实蝇幼虫为繁育寄主时,其卵历期分别为1.9、2.4与4.2 d.前裂长管茧蜂的世代发育历期与寄主龄期呈正相关关系.
前裂长管茧蜂的个体大小因寄主龄期的不同而有所差异,寄生蜂雌、雄个体大小均随寄主龄期的增加而有所增大(表2).当以1龄橘小实蝇幼虫为寄主时,所繁育出寄生蜂雌性个体的后足胫节介于1.44~1.62 mm之间,平均长度为1.52 mm,而当以2龄、3龄初及3龄末橘小实蝇幼虫为繁育寄主时,子代雌蜂个体后足胫节长度分别为1.60、1.65及1.64 mm.雄性个体也表现出相似的趋势(表2).

表1 不同龄期寄主繁育条件下前裂长管茧蜂各虫态的发育历期1)Tab.1 Developmental duration of Diachasmimorpha longicaudata reared by Bctrocera dorsalis larvae at different ages

表2 不同龄期寄主繁育条件下前裂长管茧蜂的后足胫节长度1)Tab.2 Body length of Diachasmimorpha longicaudata reared by Bctrocera dorsalis larvae at different ages
2.3 寄主龄期对前裂长管茧蜂各虫态存活率的影响
前裂长管茧蜂的个体发育与寄主龄期密切相关,寄主龄期过低或过高均对寄生蜂的生长发育产生一定的抑制作用(表3).前裂长管茧蜂各虫态的存活率均以3龄初橘小实蝇幼虫为繁育寄主时最好,卵、幼虫及蛹的存活率分别为81.22%、78.34%及62.44%,2龄的次之.从综合反映前裂长管茧蜂生长发育情况的幼期存活率来看,以3龄初橘小实蝇幼虫繁育的寄生蜂情况最好,幼期存活率为54.09%,其次为2龄的,幼期存活率为46.88%,而以1龄及3龄末橘小实蝇幼虫所繁育的寄生蜂幼期存活率较低,分别16.40%和26.29%.
2.4 寄主龄期对前裂长管茧蜂子代性比的影响
寄主年龄对前裂长管茧蜂子代性比有明显的影响.以3龄初橘小实蝇幼虫为繁育寄主时,寄生蜂子代种群中以雌性个体为主,雌性比率为64.11%.而1龄、2龄及3龄末处理中,寄生蜂子代种群则以雄性个体为主,相应的雌性比率分别为11.44%、46.57%与33.21%(图2).

表3 不同龄期橘小实蝇幼虫繁育条件下前裂长管茧蜂各虫态的存活率1)Tab.3 Survival rate of Diachasmimorpha longicaudata reared by Bctrocera dorsalis larvae at different ages
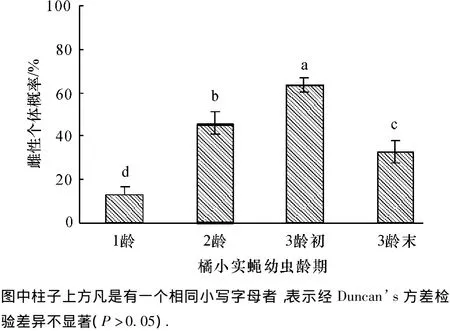
图2 不同龄期橘小实蝇幼虫繁育的前裂长管茧蜂后代中雌性个体的概率Fig.2 Percentage of female offspring of Diachasmimorpha longicaudata reared by Bctrocera dorsalis larvae at different ages
3 讨论与结论
寄主龄期是寄生蜂与寄主相互作用关系中的一个重要因素[11].寄生蜂通常倾向选择寄主生长过程中的某一阶段进行寄生,而很少攻击所有年龄段的寄主[12-14].寄生蜂对寄主龄期的选择体现出寄主的适合性问题,这种适合性往往由寄主的生理状态以及营养物质的含量决定.研究表明,寄主对寄生蜂的入侵有一定的免疫反应,且这种免疫能力与寄主生长发育阶段密切相关,通常在生长发育早期,寄主的免疫力较差,随着龄期的增加其抵抗力不断增强[11,15].另外,寄主体内营养物含量是确保顺利完成世代发育的物质基础,一般来说,随着龄期的增加,寄主所含的营养物质也逐步增加,给寄生蜂所能提供的养分也就越多[16-17].寄生蜂往往选择既能克服寄主免疫反应,同时又有足够营养物质的某一特定年龄段的寄主进行寄生,且不同类型的寄生蜂具有不同的寄主选择策略.对卵寄生蜂来说,由于寄主卵容积不会发生太大的变化,寄主的生理状态是卵寄生蜂判断寄主适合性的主要指标,如卵寄生蜂——卷蛾分索赤眼蜂喜欢寄生新鲜的(<24 h)小菜蛾卵[18].对幼虫期寄生蜂来说,除寄主的免疫能力外,寄生蜂需同时考虑寄主体内营养物质的含量,如菜蛾绒茧蜂偏好寄生3龄小菜蛾幼虫,且羽化出寄生蜂的繁殖力明显高于寄生2龄和4龄小菜蛾幼虫所羽化出的蜂[12].
从本试验的结果来看,前裂长管茧蜂对寄主龄期具有明显的选择性.在试验观察的6 h内,前裂长管茧蜂寄生2龄与3龄初的橘小实蝇幼虫的数量显著高于1龄与3龄末橘小实蝇幼虫,且具有较高的卵孵化率、幼虫与蛹的存活率高.低龄(1龄)橘小实蝇幼虫对前裂长管茧蜂的寄生与发育的影响最为显著,前裂长管茧蜂的寄生效能受到明显削弱,幼期存活率低,说明在前裂长管茧蜂的寄主选择过程中,寄主营养物质含量与寄主免疫能力相比,前者的影响可能更为突出.与此同时,寄生1龄橘小实蝇幼虫的寄生蜂卵、幼虫与蛹的发育速率加快,只需较短的时间就能完成世代发育,原因可能是当低龄橘小实蝇幼虫被寄生时,随同前裂长管茧蜂卵进入寄主体内的某些破坏寄主免疫系统的因子,如多分DNA病毒(Polydnavirus,PDA)、畸形细胞、毒液等可能抑制了寄主的生长发育,这使得寄生蜂可利用的生物资源减少,而当寄主资源缺乏时寄生蜂有可能通过提高自身的发育速率来降低后期因资源不足而导致死亡的风险[19-21].而面对高龄(3龄末)橘小实蝇幼虫时,此时寄主个体较大,体内营养物质丰富,但体内的组织、器官发育日趋完善,对寄生蜂入侵的抵抗力不断增强,寄生蜂往往难以突破寄主强大的免疫系统而导致后代死亡率上升,但一旦突破这种防御系统则有可能获得个体强大的后代.综合寄生效能、幼期存活率、子代个体大小与性比等因素来看,3龄初橘小实蝇幼虫是用于繁殖前裂长管茧蜂的最适阶段.
[1]ALUJA M,GUILLEN J,LIEDO P,et al.Fruit infesting tephritids(Dipt.:Tephritidae)and associated parasitoids in Chiapas[J],Mexico Entomophaga,1990,35:39-48.
[2]BARANOWSKY R M,GLEEN H,SIVINSKI J.Biological control of the Caribben fruit fly(Diptera:Tephritidae)[J].Fla Entomol,1993,76:245-250.
[3]LOPEZ M,ALUJA M,SLVINSKI J.Hymenopterous larval-pupal and pupal parasitoids of Anastrepha spp.(Diptera:Tephritidae)in Mexico [J].Biol Control,1999,15:119-129.
[4]HARDY D E,DELFINADO M D.Insects of Hawaii.[M]∥[Anon].A manual of the insect of the Hawaiian islands.Hawai:The University Press of Hawaii,1980:451.
[5]WONG T T Y,RAMADAN M M.Parasitization of the Mediterranean and oriental fruit fly(Diptera:Tephritidae)in the Kula area of Maui,Hawaii[J].Econom Entomol,1987,80:77-80.
[6]OVRUSKI S M,FIDALGO P.Use of parasitoids(Hym.)in the control of fruit flies(Dip.:Tephritidae)in Argentina:Bibliographic review(1937-1991)[J].IOBC/WPRS Bulletin,1994,17(6):84-92.
[7]OVRUSKI S M,ALUGA M,SIVINSKI J,et al.Hymenopteran parasitoids on fruit-infesting Tephritidae(Diptera)in Latin America and the southern United States:Diversity,distribution,taxonomic status and their use in fruit fly biological control[J].Int Peat Management Reviews,2000,5:81-107.
[8]邵屯,刘春燕,陈科伟,等.前裂长管茧蜂个体发育研究[J].环境昆虫学报,2008,30(2):147-152.
[9]邵屯,刘春燕,陈科伟,等.桔小实蝇及其寄生性天敌:前裂长管茧蜂饲养方法[J].环境昆虫学报,2008,30(4):377-380.
[10]邵屯,陈科伟,刘春燕,等.前裂长管茧蜂对桔小实蝇的寄生效能研究[J].华南农业大学学报,2009,30(2):33-36.
[11]NUSSBAUMER C,SCHOPF A.Development of the solitary larval endoparsitoid Glyptapanteles porthetriae(Hymenoptera:Braconidae)in its host Lymantria dispar(Lepidoptera:Lymantriidae)[J].Eur J Entomol,2000,97:355-361.
[12]李元喜,刘树生.寄主龄期对菜蛾绒茧蜂生物学特性的影响[J].浙江大学学报:农业与生命科学版,2001,27(1):11-14.
[13]蔡霞,郝仲萍,施祖华,等.寄主龄期对半闭弯尾姬蜂生物学特性的影响[J].中国生物防治,2006,22(2):92-95.
[14]CHAU A,MACKAUER M.Host instar selection in the aphid parasitoid Monoctonus paulensis(Hymenoptera:Braconidae,Aphidiidae):A preference for small pea aphids[J].Eur J Entomol,2000,97:347-353.
[15]李建成,张青文,刘小侠,潘文亮.中红侧沟茧蜂成虫日龄及粘虫幼虫龄期对寄生效果的影响[J].中国生物防治,2005,21(1):14-17.
[16]SEQUEIRA R,MACKAUER M.Nutritional ecology of an insect host-parasite association:The pea aphid-Aphidius ervisystem[J].Ecology,1992,73:183-189.
[17]SEQUEIRA R,MACKAUER M.The nutritional ecology of a parasitoid wasp,Ephedrus californicus Baker(Hymenoptera:Aphidiidae)[J].Can Entomol,1993,125:423-430.
[18]陈科伟,黄寿山,何余容.两种赤眼蜂对小菜蛾卵的寄生潜能分析[J].生态学报,2002,22(8):1293-1296.
[19]CAMPBELL A,MACKAUER M.The effect of parasitism by Aphidius smithi(Hymenoptera:Aphidiidae)on reproduction and population growth of the pea aphid(Homoptera:Aphididae)[J].Can Entomol,1975,107:919-926.
[20]LAWRENCE P O,MATOS L F.Transmission of the Diachasmimorpha longicaudata rhabdovirus(DIRhV)to wasp offspring:An ultrastructural analysis[J].Insect Physiology,2005,51:235-241.
[21]CYNTHIACHK,LAWRENCEP O.Hegen’s glands of the parasitic wasp Diachasmimorpha longicaudata(Hymenoptera:Braconidae):Ultrastructure and the detection of entomoposvirus and parasitism-specific protein[J].Arthropod Structure & Development,2002,31:121-130.
